DNA复制的基本规律
- DNA 以半保留复制方式进行复制:子代 DNA 中保留了亲代的全部遗传信息,亲代与子代 DNA 之间碱基序列高度一致
- DNA 复制从起点双向进行:
- 原核生物一个起点
- 真核生物多个起点,每个起点起始的 DNA 复制区称为复制子
- 双向复制在每个复制子两端形成 2 个复制叉
- DNA复制以半不连续方式进行
- DNA 聚合酶只能从 5′ → 3′ 聚合,故必然有一条链不能连续复制
- 前导链连续复制,后随链不连续复制
- 冈崎片段:后随链复制延长和解链方向不同
- DNA 复制具有高保真性
- 半保留复制
- 严格碱基配对
- 错配修复:原核生物 DNA 复制低错误率主要与 DNA pol Ⅰ 的 3’-5’ 外切酶活性(即时校对功能)有关。
- 复制叉的复杂结构:避免过早解旋导致碱基/单链DNA被修饰或水解
DNA 复制、RNA 合成(转录)、逆转录、密码阅读方向均为 5′ → 3′。
DNA 复制的酶学和拓扑学
原核生物和真核生物 DNA 聚合酶对比
| E. coli | 真核生物 | 功能 |
|---|---|---|
| Ⅰ | 去处 RNA 引物,填补复制中的 DNA 空隙,DNA 修复和重组、 对复制过程中的错误进行校对1 | |
| Ⅱ | 复制中的校对1,DNA 损伤的应急修复 | |
| β | DNA 修复 | |
| γ | 线粒体 DNA 合成 | |
| Ⅲ | ε | 前导链合成 |
| α | 引物酶 | |
| δ | 后随链合成 |
1:原核生物 DNA pol Ⅰ、Ⅱ、Ⅲ 均有校对活性(3′ → 5′ 外切酶活性),但 DNA pol Ⅱ 主要参与 SOS 修复,DNA pol Ⅲ 主要参与复制延长,发挥校对功能的主要是 DNA pol Ⅰ。
a 引物,e 前导链 → d 后随链 → c 线粒体 → b 修复原核生物 DNA 复制过程
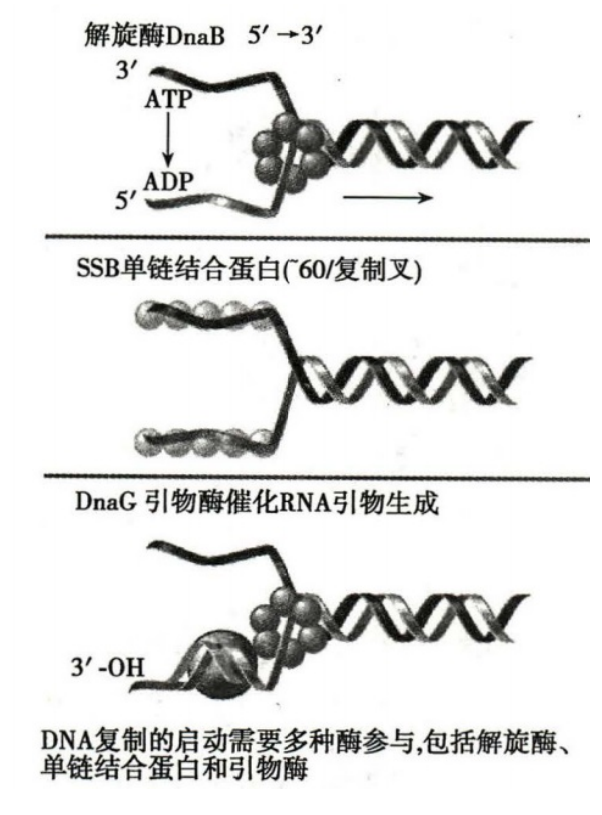
起始复合物形成
- 解松超螺旋:拓扑异构酶
- 拓扑异构酶 Ⅰ:可以切断 DNA 双链中的 1 条单链,随后又可以封闭(形成 3’,5’-磷酸二酯键,
结合连接 DNA 单链),不消耗 ATP - 拓扑异构酶 Ⅱ:可以切断 DNA 双链中的 2 条单链,随后又可以封闭,消耗 ATP
- 拓扑异构酶 Ⅰ:可以切断 DNA 双链中的 1 条单链,随后又可以封闭(形成 3’,5’-磷酸二酯键,
- DnaA:辨认、结合起始点 oriC(E.coli 上固定起点,富含 AT)
- 开链:
- DnaB(解旋酶):解开双链足够用于复制的长度,并且逐步置换出 DnaA
- DnaC:协助 DnaB,将其运到起始部位
- SSB(单链结合蛋白)和已经解开的 DNA 单链结合,稳定开链,保持复制叉的长度
- 引物合成:DnaG(引物酶)
- 引发体:引物酶进入后,DnaB、DnaC、DnaG 和复制起始点共同构成的起始复合体结构
- 引物长度一般 5–10 个核苷酸
- 利福平可以特异性的阻断 RNA 聚合酶,但是引物酶对其不敏感
- 复制的延长:DNA pol Ⅲ
- 前导链和后随链用的是同一个酶,故和真核生物有区别——没有酶的转换
- 关键亚基:
- ε 亚基:保真
- 一对 β 亚基:夹住模版链,使酶沿模版滑动
- 酶活性:5’-3’ 聚合、3’-5’ 外切(校对)
- 水解引物:DNA pol Ⅰ 大片段(Klenow 片段):5′-3′ 外切
- 填补空隙:DNA pol Ⅰ 小片段:5′-3′ 聚合、3′-5′ 外切(校对)
- 连接缺口:DNA 连接酶:
- 消耗 ATP,连接缺口
- 只能连接双链中的单链缺口,不能连接单独存在的 DNA 单链或 RNA 单链
- 是基因工程重要的工具酶之一
DNA pol Ⅱ
- DNA pol Ⅱ 基因发生突变,细菌依然能存活,推想它是在 pol I 和 pol Ill 缺失情况下暂时起作用的酶。
- DNA pol Ⅱ 对模板的特异性不高,即使在已发生损伤的 DNA 模板上,它也能催化核苷酸聚合。因此认为,它参与 DNA 损伤的应急状态修复(SOS 修复)。
真核生物 DNA 复制过程
- 复制起始和原核生物基本类似,具体过程不详。
- DNA pol α(引物)、δ(后随链)、ε(前导链) 参与复制起始
- 特点
- 每个染色体有上千个复制子,复制起点很多,复制起点较复杂
- 复制有时序性,复制子以分组方式激活,而不是同步启动
- 起点:酵母DNA复制起点含11bp富含AT的核心序列——自主复制序列(和原核生物相比有长有短)
- DNA 聚合酶:
- DNA pol α:引物酶
- DNA pol β:真核生物 DNA 修复
- DNA pol γ:真核生物线粒体 DNA 按 D-环方式复制
- DNA pol δ:后随链
- DNA pol ε:前导链
- 解旋酶
- 拓扑酶
- 复制因子:RFA、RFC
- PCNA(增殖细胞核抗原):在复制的起始和延长(和中止)中起到重要的作用
- 起始:类似于 DNA pol Ⅲ 的 β 亚基,在 RFC 的作用下结合于引物-模板(固定)
- 延长:使 DNA pol δ 获得持续合成的能力(α-δ 转换)
- 中止:促进核小体生成
- 延长过程中发生 DNA 聚合酶转换:α 和 δ 互相转换,转换频率高,催化速度慢(
真核生物复制速度慢),PCNA 全程多次发挥作用,引物、冈崎片段均比原核生物要短 - 真核生物 DNA 合成后立即组装成核小体
真核生物 DNA 复制特点
- 真核生物染色体DNA在每个细胞周期中只能复制一次
- 真核生物线粒体 DNA 按 D 环方式复制:mtDNA 为闭合环状双链结构
- 端粒酶解决染色体末端复制问题
- 意义:维持染色体稳定,DNA 复制的完整性
- 构成和功能:
- 端粒酶 RNA(hTR):提供 RNA 模版
- 端粒酶逆转录酶(hTRT):催化逆转录(RNA 指导的 DNA 聚合酶活性)
- 端粒酶协同蛋白 1(hTP1)
- 过程:
- 端粒酶逆转录酶通过端粒自身的 RNA 模板,指导延长 DNA 母链
- 引物酶:RNA 引物酶通过新合成的母链,合成引物)
- 招募 DNA pol,以 DNA 母链为模板,沿着新合成的引物进行填充
- 水解 RNA 引物
逆转录
- 引物:逆转录的引物由病毒颗粒自身的 tRNA 提供。
- 逆转录酶活性:
- RNA 指导的 DNA 聚合酶活性(RNA → DNA):RNA 指导的 DNA 聚合
- RNase H(Hybrid)活性:水解杂化双链中的 RNA 单链
- DNA 指导的 DNA 聚合酶活性(DNA → DNA):DNA 指导的 DNA 聚合(没有校对活性)“DNA 模板也可直接用于/参与逆转录”
- 逆转录的意义:
- 遗传信息从 RNA 流动到 DNA
- 发展了中心法则
- 拓宽了病毒致癌理论,从逆转录病毒中发现了癌基因(HIV 也是 RNA 病毒,有逆转录活性)
- 是获取目的基因重要的方法之一——cDNA 法
DNA 损伤
DNA 损伤的原因与DNA 损伤之间为复杂的多对多关系,掌握嘧啶二聚体即可:
嘧啶二聚体
- 原因:紫外照射(
电离辐射) - 损伤类型:DNA 链共价交联(
碱基损伤) - 修复方式:直接修复(DNA 光裂合酶-光修复)
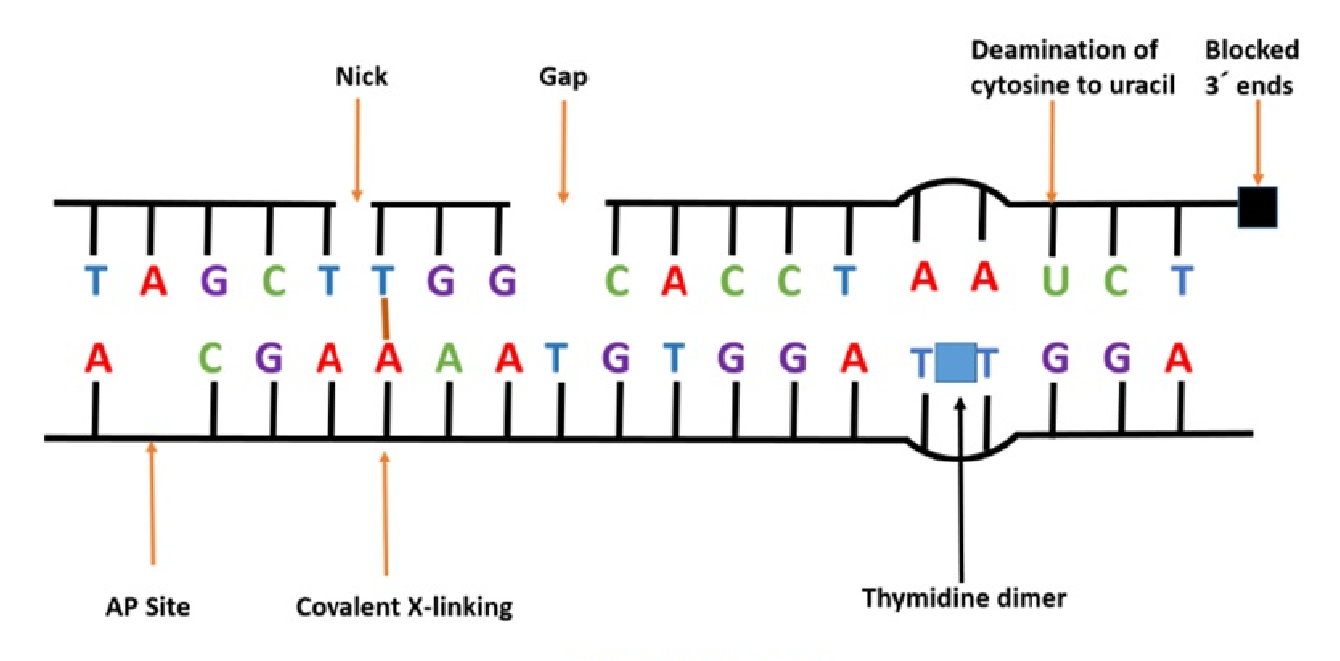
DNA 损伤类型
- 损伤的类型:碱基损伤与糖基破坏,碱基之间发生错配,DNA 链发生断裂(电离辐射造成 DNA 损伤的主要形式),链内/链间/DNA-蛋白质,DNA 链的共价交联(嘧啶二聚体)
- 损伤的结果:可导致 DNA 模板发生碱基置换、插入、缺失、链的断裂等变化,并影响染色体的高级结构
- 碱基置换
- 转换:嘌呤换嘌呤或者嘧啶换嘧啶
- 颠换:嘌呤和嘧啶之间互换
- 可引起碱基错配,导致基因突变
- 可能但是不一定引起氨基酸编码的改变(简并性)
- 错义突变:改变氨基酸编码
- 同义突变:不改变氨基酸编码
- 无义突变:变为终止密码子
- 碱基插入和缺失:移码突变
- DNA 断裂:阻止 RNA 合成过程中链的延伸
DNA 损伤修复
- 直接修复
- 嘧啶二聚体的直接修复:DNA 光裂合酶-光修复
- 烷基化碱基的直接修复:烷基转移酶
- 切除修复:最普遍的 DNA 损伤修复方式
- 碱基切除修复:
- 识别水解:DNA 糖苷酶,识别受损碱基,去除后形成无碱基位点(AP 位点)
- 切除:无碱基位点核酸内切酶,切掉剩下的磷酸核糖
- 合成:DNA 聚合酶合成互补序列
- 连接:DNA 连接酶将切口重新连接
- 核苷酸切除修复:
- 识别损伤对 DNA 双螺旋结构所造成的扭曲
- 损伤部位两侧切开 DNA 链,去除切口间的寡核苷酸
- DNA 聚合酶作用下,填补缺损
- DNA 连接酶连接缺口
- 碱基错配修复:是碱基切除修复的一种特殊形式
- 碱基切除修复:
- 重组修复:双链断裂的修复形式
- 同源重组修复
- 非同源末端连接的重组修复
- 跨越损伤 DNA 合成:应用于 DNA 双链发生大范围损伤时,跨越过损伤部位进行复制,随后再修复
- 重组跨越损伤修复:拆东墙补西墙,损伤没有被真正的修复,转移到了另一个 DNA 分子上
- 合成跨越损伤修复:
- SOS 修复
- DNA pol Ⅳ、Ⅴ
- 活性低,识别碱基精确度差,无校对功能,故出错的概率较大
- 产生新的聚合酶(DNA pol Ⅳ、Ⅴ)替代原来的 DNA 聚合酶 Ⅲ
- 随机插入核苷酸进行合成
- 跨越损伤后,新的 DNA 聚合酶从 DNA 链上脱离,再由原来的 DNA 聚合酶 Ⅲ 继续复制
- DNA pol Ⅱ 也参与 SOS 修复,但需要校对
- DNA pol Ⅳ、Ⅴ
- SOS 修复
DNA 损伤修复的意义
- DNA 损伤具有双重效应
- 可造成 DNA 突变,突变是进化的分子基础
- 可造成细胞功能障碍,甚至死亡
- DNA 损伤修复障碍与多种疾病相关 简单看看
- 着色性干皮病:核苷酸切除修复
- 遗传性非息肉性结肠癌:错配修复、转录偶联修复(DNA 修复调节基因)
- 遗传性乳腺癌:同源重组修复
- Bloom综合征:非同源末端连接重组修复
- 范可尼贫血:重组跨越损伤修复
- Cockyne综合征:核苷酸切除修复、转录偶联修复
- 毛发低硫营养不良:核苷酸切除修复