神经元和神经胶质细胞
神经元
神经元的一般结构
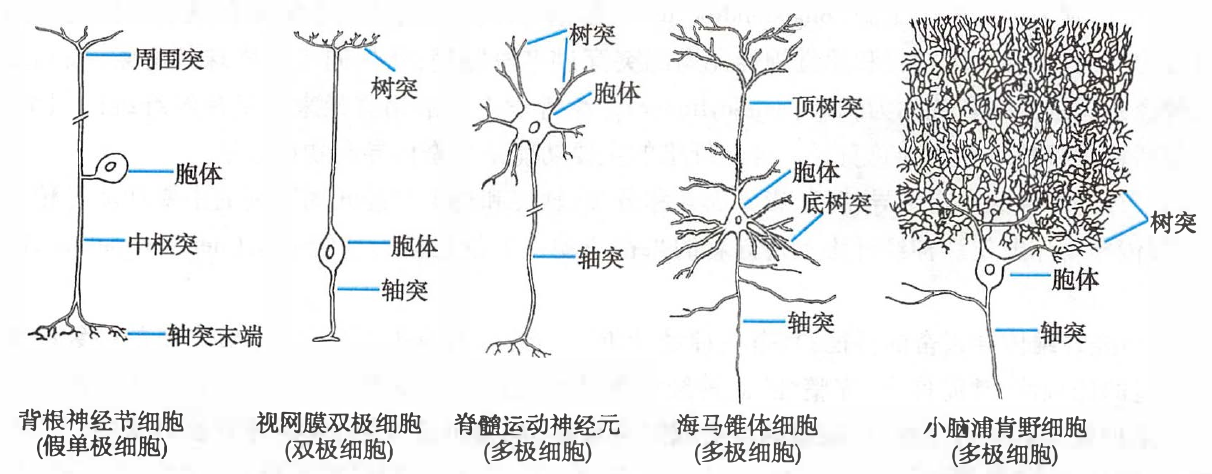
哺乳动物神经系统中几种不同类型的神经元模式图
- 树突:
- 与其他神经元的轴突末梢形成突触
- 在大脑皮层,约 98% 的突触由树突参与形成,仅约 2% 由胞体参与形成
- 树突的分支及树突棘都使细胞膜面积大幅扩展,从而提高了神经元信息接收的范围和敏感性
- 在脑发育期,树突棘数量的不断增加与智力的发育有关
- 轴突:
- 通常只有一条
- 轴丘:胞体发出轴突的部位膨大并向外突起
- 轴突始段一般略为粗大,且无髓鞘包裹
- 轴突末段:
- 在一个神经元与另一个神经元或效应细胞相接触而形成的突触结构中,轴突末端通常构成突触前部分
- 轴突末段分成许多分支,完全无髓鞘包裹,称为神经末梢,其最末端常膨大为球状、纽扣状或柄状,称为突触小扣、终扣或突触小结
树突和轴突在结构上为神经元赋予了区域性或极性,为神经元的区域性功能分化提供了结构基础,也为神经元形态分类提供了依据。
神经元的主要功能
神经元的主要功能是接受、整合、传导和传递信息(传导兴奋,传导兴奋是神经纤维的功能):
- 胞体和树突:主要负责接受和整合信息;
- 轴突始段:主要负责产生动作电位,也参与信息整合;
- 轴突:负责传导信息;
- 突触末梢:负责向效应细胞或其他神经元传递信息。
神经纤维及其功能
轴突和感觉神经元的周围突都称为神经纤维,其主要功能是兴奋传导和物质运输。
神经纤维的分类
| Erlanger-Gasser 分类(传出) | 对应的 Lloyd-Hunt 分类(传出) | 功能 |
|---|---|---|
A(有髓鞘) | ||
| α | Ⅰa、Ⅰb | 本体感觉、躯体运动 |
| β | Ⅱ | 触压觉 |
| γ | 支配梭内肌(引起收缩) | |
| δ | Ⅲ | 痛觉、温度觉、触-压觉(快) |
B(有髓鞘) | 自主神经节前纤维 | |
C(无髓鞘) | ||
| 后根 | Ⅳ | 痛觉、温度觉、触-压觉(慢) |
| 交感 | 交感节后纤维 |
- 髓鞘:
- 构成髓鞘或神经膜的胶质细胞
- 在周围神经系统主要是施万细胞
- 在中枢为少突胶质细胞
- 髓鞘的厚度往往远大于轴突和感觉神经元的周围突自身的直径
- 构成髓鞘或神经膜的胶质细胞
- 轴索:当轴突和感觉神经元周围突穿过多段髓鞘,就如同串起串珠的绳索,故轴突和感觉神经元周围突两者统称为轴索
神经纤维的兴奋传导功能
轴突和感觉神经元周围突(神经纤维)的主要功能是传导兴奋,亦即传导动作电位。神经纤维上传导着的兴奋或动作电位也称为神经冲动(nerve impulse),简称冲动。
- 完整性(对完整的神经纤维结构和功能的依赖性):神经纤维只有在其结构和功能都完整的情况下才能传导兴奋。
- 绝缘性(互不干扰性):一条神经干内含多条神经纤维,但因为被相对大容量的细胞外液“浸泡”,且单条神经纤维上传导神经冲动的局部电流又很微弱,多条神经纤维同时传导兴奋时互不干扰。
- 双向性:在神经纤维的一个局部发生的动作电位,(在离体情况下)会同时向相反的两个方向传导。
- 多数轴突总是将神经冲动由胞体传向末梢
- 但作为轴突的感觉神经周围突则将神经冲动传向胞体
- 相对不疲劳性:(相对突触而言)神经纤维能长时间保待其传导兴奋的能力。
- 直径:一般来说,神经纤维直径愈大,传导速度越快
- 髓鞘有无:有髓神经纤维传导速度比无髓神经纤维快
- 髓鞘厚度:在一定范围内,有髓神经纤维的髓鞘愈厚,传导愈快
- 温度
神经纤维的轴浆运输功能
- 顺向轴浆运输:自胞体向轴突末端
- 快速顺向轴浆运输(由驱动蛋白执行):主要见于具有膜结构的细胞器,如线粒体、突触毅泡和分泌颗粒等的运输。
- 慢速轴浆运输:新在胞体合成以及刚从微管和微丝的分解端解离下来的微管蛋白、神经微丝蛋白等细胞骨架成分。
- 逆向轴浆运输(由动力蛋白及其辅助因子执行):自末梢到胞体主要见于某些被轴突末梢摄取的物质,如神经营养因子、狂犬病病毒、破伤风毒素等的运输。
神经对效应组织的营养性作用
- 神经通过末梢释放神经递质引起所支配的组织迅速执行其主要功能,如肌肉收缩、腺体分泌等,称为神经的功能性作用。
- 神经末梢还释放某些营养因子,调整所支配组织的代谢活动,缓慢但持续地影响其结构和功能状态,这类作用称为神经的营养性作用。
脊髓灰质炎患者的肌肉萎缩,即主要因支配相应肌肉的脊髓中央灰质前角运动神经元变性死亡,对肌肉失去营养作用所致。
神经胶质细胞
- 胶质细胞的结构和功能特征
- 胶质细胞也有突起,但无树突和轴突之分
- 细胞之间不形成化学性突触,但普遍存在缝隙连接(神经元之间也存在缝隙连接,即电突触)
- 膜电位也随细胞外 K+ 浓度而改变,但不能产生动作电位
- 某些胶质细胞膜上还存在多种神经递质的受体
- 终身具有分裂增殖的能力
- 胶质细胞的类型和功能
- 星形胶质细胞:机械支持和营养作用;隔离和屏障作用;迁移引导作用;修复和增生作用;免疫应答作用;细胞外液中 K+ 浓度稳定作用(受损可导致局部癫痫病灶形成);对某些递质和活性物质的代谢作用
- 少突胶质细胞和施万细胞:
- 结构:分别在中枢(少突胶质细胞)和周围(施万细胞)神经系统形成髓鞘
- 功能:
- 在有髓的神经纤维,髓鞘使动作电位跳跃式传导,可大大提高神经纤维传导兴奋速度
- 髓鞘能引导轴突生长并促进其与其他细胞建立突触联系
- 在周围神经损伤变性后的再生过程中,轴突可沿施万细胞所构成的索道生长
- 其他各类胶质细胞:
- 小胶质细胞:
- 相当于中枢神经系统中的吞噬细胞
- 脑组织发生变性时,小胶质细胞能转变成巨噬细胞,与来自血液中的单核细胞和血管壁上的巨噬细胞一起清除变性的神经组织碎片
- 脉络丛上皮细胞和室管膜细胞:作为胶质细胞,通过形成紧密连接参与血-脑脊液屏障和脑-脑脊液屏障的构成
- 卫星细胞:
- 存在于周围神经系统的脊神经节内
- 其作用可能是为神经元提供营养及形态支持,以及调节神经元外部的化学环境
- 小胶质细胞:
突触传递
电突触传递
电突触是以电流为传递媒质的突触,其结构基础是缝隙连接,具有双向性、快速性、低电阻等特点。
化学性突触传递
根据突触前、后两部分之间有无紧密的解剖学关系,可将化学性突触分为定向突触和非定向突触。
定向突触传递
定向突触末梢释放的递质仅作用于突触后范围极为局限的部分。
定向突触的结构
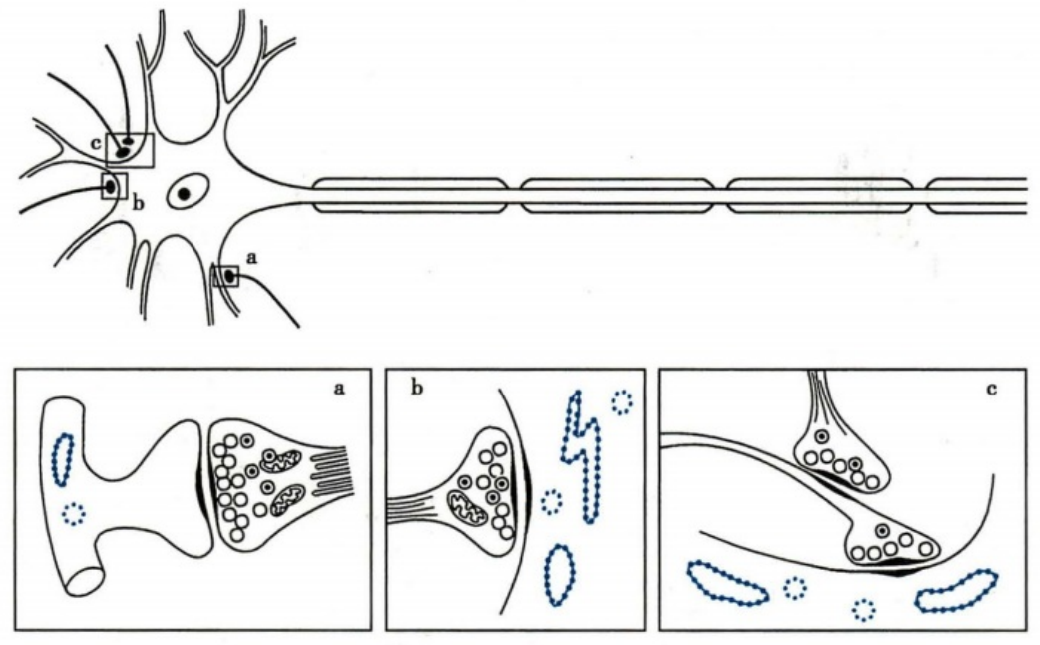
突触联系基本类型模式图
a、b、c 分别为轴突-树突式、轴突-胞体式、轴突-轴突式联系
- 突触前膜:突触前末梢的轴浆内有密集的线粒体和突触囊泡
- 活化区:突触囊泡密集
- 突触间隙:内含高浓度神经递质
- 突触后膜:密集分布着特异性受体或递质门控通道
- 突触后致密区:聚集着大量细胞骨架和信号蛋白分子
- 小而清亮透明的囊泡:内含乙酰胆碱或氨基酸类递质;
- 小而具有致密中心的囊泡:内含儿茶酚胺类递质;
- 大而具有致密中心的囊泡:内含神经肽类递质。
经典突触的传递过程
- 突触前神经元的兴奋传到末梢时,突触前膜去极化
- 去极化达一定程度时,前膜上的电压门控钙通道开放,Ca2+ 内流
- 轴浆内 Ca2+ 浓度迅速升高,触发突触囊泡的出胞
- 递质的释放量与进入轴浆内的 Ca2+ 量呈正相关
- 这一过程结束后,轴浆内积聚的 Ca2+ 主要经由 Na+-Ca2+ 反向转运体(交换体)迅速被转运到细胞外
- 神经递质以囊泡为单位释放的方式称为量子释放
- 神经递质释放机制:动员(钙调蛋白)、摆渡(小 G 蛋白 Rab3)、着位、融合和出胞
- 神经递质释入突触间隙后,经扩散抵达突触后膜并作用于其上的特异性受体或递质门控通道
- 后膜对某些离子通透性的改变,使某些带电离子进出后膜,或使进出量发生改变
- 突触后膜发生一定程度的去极化或超极化的电位变化(突触后电位)
影响定向突触传递的因素、环节
- 影响递质释放的因素:递质的释放量主要决定于进入末梢的 Ca2+ 量,凡能影响末梢处 Ca2+ 内流的因素都能改变递质的释放量
- 使递质释放增多:细胞外 Ca2+ 浓度升高、到达突触前末梢动作电位的频率加快、幅度增加或时程延长等。
- 使递质释放减少:细胞外 Mg2+ 浓度升高、钙通道密度降低、各种钙通道桔抗剂、一些神经毒素等。
- 突触前受体:突触前膜上还存在突触前受体,它们可在某些神经递质或调质的作用下改变进入末梢的 Ca2+ 量,进而影响递质的释放量
- 神经毒素:一些神经毒素也通过影响递质释放起作用,如肉毒梭菌毒素、破伤风毒素、毒蛛毒素
- 影响递质清除的因素:凡能影响递质重摄取和酶解代谢的因素也能影响突触传递
- 三环类抗抑郁药:可抑制脑内 NE 在突触前膜的重摄取,使递质滞留于突触间隙而持续作用于受体,从而使传递效率加强。
- 利血平:能抑制囊泡膜对 NE 的重摄取,使 NE 滞留在末梢轴浆内而被单胺氧化酶酶解,囊泡内递质减少以至耗竭,可用于缓解舞蹈病症状。
- 新斯的明及有机磷农药等可抑制突触后膜上的乙酰胆碱酯酶,阻碍乙酰胆碱水解使其持续发挥作用。
- 影响突触后膜反应性的因素:
- 受体发生上调或下调:递质释放量发生改变时,突触后受体的密度及与递质结合的亲和力均可发生改变
- 突触后膜受体的阻断剂:筒箭毒碱和 α-银环蛇毒可阻断胆碱能突触后膜的 N2 型乙酰胆碱受体通道。
- 临床上使用筒箭毒碱等作为肌松剂。
非定向突触传递非突触性化学传递
非定向突触传递不具有经典突触的结构,其突触前末梢释放的递质可扩散至距离较远和范围较广的突触后成分,所以也称为非突触性化学传递。
- 在中枢神经系统:主要发生于单胺能(肾上腺素能、多巴胺能及 5-羟色胺能)神经元的纤维末梢部位
- 在周围神经系统:自主神经节后纤维(主要是交感神经节后纤维)与效应细胞之间的接头
- 曲张体
- 内有大量突触囊泡的膨大结构
- 不与突触后效应细胞形成经典的突触联系,而是随分支抵达效应细胞的近旁
- 特点
- 无特定的突触后成分,因而作用部位较分散
- 无固定的突触间隙,因而递质扩散的距离远近不等,时间长短不一,曲张体与效应器之间的距离一般大于 20 nm,有的甚至超过 400 nm
- 释放的递质能否产生信息传递效应,取决于靶细胞上有无相应的受体
- 曲张体
突触后电位
突触前膜释放的神经递质作用于突触后膜上的相应受体后,会引起突触后膜去极化或超极化的突触后电位(均为局部电位)(包括接头电位),亦即引起突触后神经元的兴奋性变化。
IPSP 的产生还可能与突触后膜钾通道的开放或钠通道和钙通道的关闭有关
- 兴奋性突触后电位(EPSP):突触传递在突触后膜引起的去极化突触后电位。
- 快 EPSE:
- 举例:来自伸肌肌梭的传入冲动在脊髓前角伸肌运动神经元引起的去极化
- 机制:兴奋性递质作用于突触后膜的相应受体,使某些离子通道开放,后膜对 Na+ 和 K+ 的通透性增大,但因 Na+ 内流大于 K+ 外流,故发生净内向电流,导致后膜出现去极化。
- 慢 EPSP:
- 机制:多与 K+ 电导降低有关
- 举例:部分交感神经节后神经元、大脑皮层神经元(教材未述)
- 举例:
- 快 EPSE:
- 抑制性突触后电位(IPSP):突触传递在突触后膜引起的超极化突触后电位。
- 快 IPSP:
- 机制:是抑制性中间神经元释放的抑制性递质作用于突触后膜,使后膜上的氯通道开放,引起外向电流(Cl- 内流),结果使突触后膜发生超极化。 IPSP 的产生还可能与突触后膜钾通道的开放或钠通道和钙通道的关闭有关。
- 举例:来自伸肌肌梭的传入冲动通过抑制性中间神经元在与该伸肌相拮抗的屈肌的运动神经元所引起的超极化
- 慢 IPSP:
- 机制:钾通道的开放
- 快 IPSP:
突触后神经元动作电位的产生
- 一个突触后神经元一般与多个突触前神经末梢构成突触,既产生 EPSP 也产生 IPSP,突触后神经元胞体电位改变的总趋势取决于同时或几乎同时产生的 EPSP 和 IPSP 的总和。
- 当其膜电位总趋势为超极化时,突触后神经元表现为被抑制
- 当其膜电位总趋势为去极化时,则易于达到阈电位而爆发动作电位,即兴奋性提高
- 起始部位:
- 动作电位首先发生在轴突始段,这是因为电压门控钠通道在该段轴突膜上密度较大,而在胞体和树突膜上则很少分布。
- 在感觉神经元,动作电位可爆发于其有髓周围突远端的第一个朗飞结处,或无髓周围突远端的未明确部位,然后向胞体方向传导。
- 动作电位一旦爆发,既可沿轴突传向末梢,也可逆向传到胞体
- 逆向传导的意义可能在于消除神经元此次兴奋前不同程度的去极化或超极化的影响,使其状态得到一次“重启”
神经递质和受体
神经递质
神经递质是指由突触前神经元合成并释放,能特异性地作用于突触后神经元或效应细胞上的受体而产生一定效应的信息传递物质,是神经元之间或神经元与效应细胞间主要的信息传递物质。
递质的鉴定
- 突触前神经元具有合成递质的前体和酶系统,并能合成该递质
- 合成后的递质贮存于突触囊泡内,并能在兴奋冲动抵达末梢时释放入突触间隙
- 能作用于突触后膜上的特异受体并发挥其生理作用,人为给突触后神经元或效应细胞施用递质,应能引起相同的生理效应
- 存在使该递质失活的酶或其他失活方式(如重摄取)
- 存在能分别模拟或阻断该递质突触传递作用的特异性受体激动剂和拮抗剂
- 有些物质(如一氧化氮、一氧化碳等)虽不完全符合上述经典递质的 5 个条件,但所起的作用与递质完全相同,故也将它们视为神经递质
神经递质的分类
- 胆碱类:乙酰胆碱
- 胺类:去甲肾上腺素、肾上腺素、多巴胺、5-羟色胺、组胺
- 氨基酸类:谷氨酸、门冬氨酸、γ-氨基丁酸、甘氨酸
- 肽类:P 物质和其他速激肽、阿片肽、下丘脑调节肽、血管升压素、缩宫素、脑-肠肽、钠尿肽、降钙素基因相关肽、神经肽 Y 等
- 嘌呤类:腺苷、ATP
- 气体类:一氧化氮、一氧化碳
- 脂类:花生四烯酸及其衍生物(前列腺素等)、神经活性类固醇
神经调质
对递质信息传递起调节作用的物质称为神经调质。
- 实际上,递质和调质之间有时并无十分明确的界限
- 神经调质的来源并不限于神经元,有些内(旁)分泌激素、免疫系统的信使物质对突触传递也有调制作用,也被归于神经调质范畴
传出神经系统分类
传出神经系统包括植物神经系统和运动神经系统,前者又称为自主神经系统,分为交感神经和副交感神经。
传出神经根据末端释放的递质不同,分为
- 胆碱能神经:以乙酰胆碱为递质,包括
- 交感神经的所有节前纤维:如支配肾上腺髓质的内脏大神经;
- 交感神经的少数节后纤维:支配多数小汗腺的纤维和支配骨骼肌血管的舒血管纤维;
- 副交感神经的大部分节前、节后纤维(少数副交感节后纤维释放肽类和嘌呤类递质);
- 运动神经:支配骨骼肌。
- 去甲肾上腺素能神经:主要以去甲肾上腺素为递质,包括交感神经的大多数节后纤维。
由于大多数交感神经节后纤维节后纤维的递质为去甲肾上腺素,所以除了促进汗腺分泌和骨骼肌血管收缩外,交感神经的功能几乎都与去甲肾上腺素的作用一致。
递质共存
两种或两种以上的递质(包括调质)共存于同一神经元内,这种现象称为递质共存。
- 生理意义:协调某些生理功能活动
- 举例:猫唾液腺接受副交感神经和交感神经的双重支配
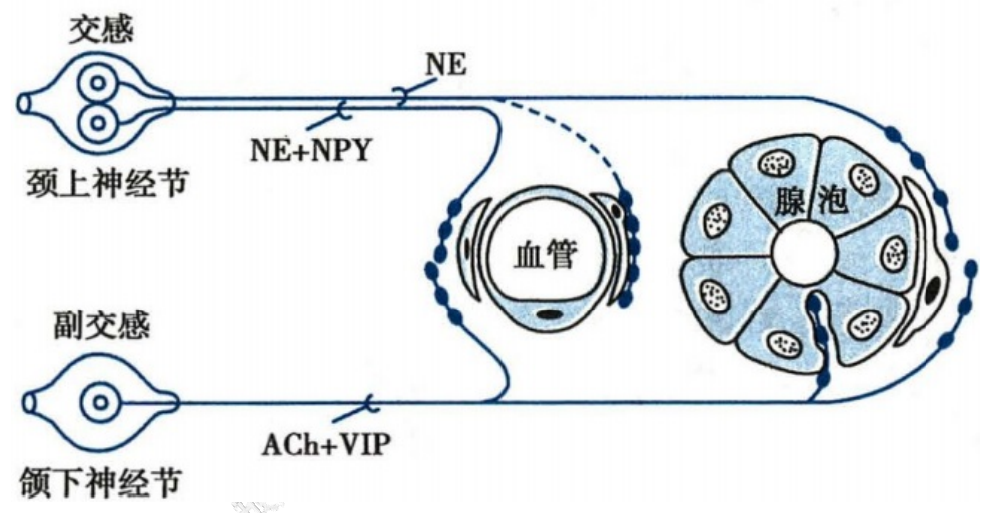
猫唾液腺递质共存示意图
- 副交感神经
- 内含乙酰胆碱:能引起唾液分泌
- 血管活性肠肽:可舒张血管,增加唾液腺的血供,并增强唾液腺上胆碱能受体的亲和力
- 交感神经
- NE:促进唾液分泌和减少血供的作用
- 神经肽 Y:主要收缩血管,减少血供
- 副交感神经
受体
受体是指位于细胞膜上或细胞内能与某些化学物质(如递质、调质、激素等)特异结合并诱发特定生物效应的特殊生物分子。
乙酰胆碱及其受体
以 ACh 为递质的神经元称为胆碱能神经元,其神经纤维称为胆碱能纤维,能与 ACh 特异性结合的受体称为胆碱能受体。
胆碱能纤维和受体的分布
- 胆碱能纤维的分布
- 中枢:
- 脊髓前角运动神经元和丘脑后部腹侧的特异性感觉投射神经元
- 脑干网状结构上行激动系统的多个环节、纹状体、前脑基底核、边缘系统的梨状区、杏仁核、海马等部位的部分神经元
- 外周:
- 自主(交感、副交感)神经节前纤维(后膜有 N1 受体)
- 大多数副交感节后纤维(少数释放肽类或嘌呤类递质的纤维除外)
- 少数交感节后纤维:如支配多数小汗腺的纤维和支配骨骼肌血管的舒血管纤维
- 骨骼肌运动神经纤维(后膜有 N2 受体)
- 中枢:
- 胆碱能受体的分布:
- M 受体(毒蕈碱受体)(从上到下分布:脑、心、平滑肌、胰腺):为 G 蛋白耦联受体
- 脑内:
- M1 受体在脑内含量丰富
- 外周:分布于大多数副交感节后纤维支配的效应细胞、汗腺细胞、骨骼肌血管的平滑肌细胞
- M2 受体主要分布于心脏
- M3、M4 受体存在多种平滑肌
- M4 受体还见于胰腺腺泡和胰岛组织,介导胰酶和胰岛素的分泌
- M5 受体的情况不详
- 脑内:
- N 受体(烟碱受体):是促离子型受体,具有递质门控特性
- N1 型(神经元型):中枢神经系统、自主神经节后神经元
- N2 型(肌肉型):骨骼肌神经-肌接头处的终板膜
- M 受体(毒蕈碱受体)(从上到下分布:脑、心、平滑肌、胰腺):为 G 蛋白耦联受体
胆碱能受体的效应及阻断剂
中枢神经系统的几乎所有功能都有胆碱能系统参与;周围胆碱能系统主要涉及自主神经系统和骨骼肌活动的调节。
- 毒蕈碱样作用(M 样作用):
- 烟碱样作用(N 样作用):
- 效果:
- 小剂量:兴奋自主神经节节后神经元;收缩骨骼肌。
- 大剂量:阻滞自主神经节。
- 阻断剂:
- 筒箭毒碱:N 受体阻断剂
- 六烃季铵、美加明:选择性 N1 受体拮抗剂,用于控制严重高血压。
- 十烃季铵、戈拉碘铵:选择性 N2 受体拮抗剂,用作肌松药。
- 效果:
- 虹膜环形肌/瞳孔括约肌,收缩时瞳孔缩小
- 虹膜辐射状肌/瞳孔开大肌,收缩时瞳孔扩大
单胺类递质及其受体
- 神经元胞体在中枢分布相对集中
- 纤维投射及受体分布的范围非常广泛
- NE能神经元胞体绝大多数位于低位脑干,尤其是中脑网状结构、脑桥的蓝斑以及延髓网状结构的腹外侧部分
- 多巴胺能神经元胞体主要集中在中脑黑质
- 5-羟色胺能神经元胞体主要集中于低位脑干的中缝核内
- 组胺能神经元的胞体集中于下丘脑后部的结节乳头核内
去甲肾上腺素和肾上腺素及其受体
肾上腺素能纤维
以 NE 为递质的神经元称为 NE 能神经元,以肾上腺素为递质的神经元称为肾上腺素能神经元,以肾上腺素或 NE 为递质的神经纤维均称为肾上腺素能纤维。
- NE 能神经元:
- *中枢:主要位于低位脑干部分,尤其是中脑网状结构、脑桥蓝斑、延髓网状结构的腹外侧区
- 上行部分:投射到大脑皮层、边缘前脑和下丘脑
- 下行部分:投射至脊髓后角的胶质区、侧角和前角
- 外周:NE 是多数交感节后纤维(除支配汗腺和骨骼肌血管的交感胆碱能纤维外)释放的递质
- *中枢:主要位于低位脑干部分,尤其是中脑网状结构、脑桥蓝斑、延髓网状结构的腹外侧区
- 肾上腺素能神经元和肾上腺素能纤维:
- 目前仅见于中枢神经系统内,其胞体主要分布在延髓,纤维投射也有上行和下行部分
- 肾上腺素在外周仅作为一种内分泌激素,由肾上腺髓质合成和分泌
- 所谓外周肾上腺素能纤维,其递质实际上是 NE
肾上腺素能受体
能与 NE 和肾上腺素结合的受体称为肾上腺素能受体,均属 G 蛋白藕联受体。
- α 受体:
- α1 受体(兴奋):血管(皮肤、肾脏、胃肠道血管 α1 受体占优势)、子宫、虹膜辐射状肌(瞳孔开大肌)
- α2 受体(抑制):小肠运动减弱
- β 受体:
- β1 受体(兴奋):心肌
- β2 受体(抑制):血管(骨骼肌、肝血管 β2 受体占优势)、子宫、小肠、支气管
- β3 受体:主要分布于脂肪组织,与脂肪分解有关
糖原合成(胰岛素释放)与 α2、β2 受体有关,糖原分解(胰高血糖素释放)与 β2 受体有关。
- 肾上腺素在外周作为内分泌激素,对 α 和 β 受体的作用都很强。
- 对血管的作用:
- 小剂量:以兴奋 β2 受体为主,血管平滑肌舒张。
- 大剂量:α1 受体也兴奋,引起血管收缩。
- 对血管的作用:
- NE 对 α 受体的作用较强,而对 β 受体的作用较弱(β1 > β2)。
判断某器官上分布的受体思路
- 判断交感兴奋时,该器官的效应是兴奋还是抑制,如果是兴奋则写 1,抑制则写 2
- 如果是兴奋:只要不是心脏(β1),都是 α1
- 如果是抑制:只要不是小肠(α2),都是 β2
多巴胺及其受体
中枢 DA 系统主要存在于中枢黑质-纹状体、中脑-边缘前脑、结节-漏斗三条通路,分别与运动调控、奖赏行为和成瘾、垂体内分泌活动调节等有关。
受体的调节
- 当递质分泌不足时,受体的数量将逐渐增加,亲和力也逐渐升高,称为受体的上调;反之则称为受体的下调
- 由于膜的流动性,储存于胞内膜结构上的受体蛋白可通过胞吐融合于细胞膜上,使发挥作用的受体数量增多
- 细胞膜上的受体也可通过受体蛋白的内吞入胞,即内化,减少膜上发挥作用的受体数量
反射活动的基本规律
反射的分类
- 非条件反射:
- 是指生来就有、数量有限、比较固定和形式低级的反射活动,如防御反射、食物反射、性反射等
- 非条件反射是人和动物在长期的种系发展中形成的
- 它的建立无需大脑皮层的参与,通过皮层下各级中枢就能形成
- 它使人和动物能够初步适应环境,对于个体生存和种系生存具有重要意义
- 条件反射:
- 是指通过后天学习和训练而形成的反射
- 它是反射活动的高级形式,是人和动物在个体生活过程中按照所处的生活环境,在非条件反射的基础上不断建立起来的,其类型和数量并无定数,可以建立,也能消退
- 人和高等动物形成条件反射的主要中枢部位在大脑皮层
- 与非条件反射相比,条件反射使人和高等动物对各种环境具有更加完善的适应性
反射的中枢整合
- 单突触反射:
- 传入神经元和传出神经元之间,在中枢只经过一次突触传递的反射
- 腱反射(如膝反射、肱二头肌反射等)是体内唯一仅通过单突触反射即可完成的反射
- 多突触反射
- 在中枢经过多次突触传递的反射
- 人和高等动物体内的大部分反射都属于多突触反射
- 整合:
- 在整体情况下,无论是简单的还是复杂的反射,传入冲动进入脊髓或脑干后,除在同一水平与传出部分发生联系并发出传出冲动外,还有上行冲动传到更高级的中枢部位进一步整合,再由高级中枢发出下行冲动来调整反射的传出冲动
- 进行反射时,既有初级水平的整合,也有较高级水平的整合
- 在通过多级水平的整合后,反射活动变得更复杂和更具适应性
中枢神经元之间的联系方式
- 单线式联系(一对一):是指一个突触前神经元仅与一个突触后神经元发生突触联系。
- 举例:如视网膜视锥系统(中央凹处)的联系方式,单线式联系使视锥系统具有较高的分辨能力。
- 绝对的单线式联系其实很少见,会聚程度较低的突触联系也通常被视为单线式联系。
- 辐散和聚合式联系
- 辐散式联系(一对多):是指一个神经元通过其轴突侧支或末梢分支与多个神经元形成突触联系
- 在传入通路中较多见。
- 举例:如在脊髓中央灰质后角,传入神经元既有纤维分支与本节段脊髓的中间神经元及传出神经元发生联系,又有上升与下降的分支在邻近或远隔的脊髓节段与中间神经元发生突触联系
- 聚合式联系(多对一):
- 在传出通路中较多见。
- 举例:如脊髓中央灰质前角运动神经元接受不同轴突来源的突触传入。
- 辐散式联系(一对多):是指一个神经元通过其轴突侧支或末梢分支与多个神经元形成突触联系
- 链锁式和环式联系:在神经通路中,若由中间神经元构成的辐散与聚合式联系同时存在,则可形成链锁式联系或环式联系。
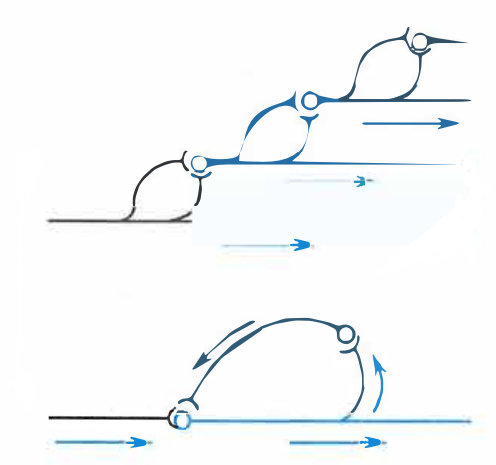
链锁式(上)和环式(下)联系
- 链锁式联系:可扩大空间作用范围。
- 环式联系:
- 特征是后一级的神经元会通过其侧支再次与前一级神经元发生突触联系,从而在结构和功能联系上都形成闭合的环路。
- 可因负反馈而使活动及时终止,也可因正反馈而使兴奋增强和延续。
- 在环式联系中,即使最初的刺激已经停止,传出通路上的冲动发放仍能继续一段时间,这种现象称为后发放或后放电。
中枢兴奋传播的特征化学性突传递的特征
- 单向传播:在反射活动中,兴奋经化学性突触传递,只能从突触前末梢传向突触后神经元。
- 中枢延搁:在一个反射活动中,从施加刺激到出现反应的时间,称为反应时间。所以从反应时间中减去兴奋在传入与传出途中所需的传导时间以及兴奋在效应器突触传递所需的时间,剩余的时间即为中枢延搁。
- 兴奋的总和:单根纤维单个传入冲动引起的 EPSP 是局部电位,其去极化幅度较小,明显小于骨骼肌单个终板电位,一般不能引发突触后神经元出现动作电位;但若干传入纤维引起的多个 EPSP 发生空间与时间总和,则容易达到阀电位水平而爆发动作电位。
- 兴奋节律的改变:反射过程中某一反射弧的传入神经(突触前神经元)和传出神经(突触后神经元)在兴奋传递过程中的放电频率往往不同,最后传出冲动的频率取决于各种影响因素的综合效应。
- 后发放与反馈:后发放见中枢神经元之间的联系方式。
- 对内环境变化敏感:突触间隙与细胞外液相通,易受内环境理化因素的变化的影响。
- 易疲劳:
- 用高频电脉冲长时间连续刺激突触前神经元,突触后神经元的放电频率将逐渐降低(如肌肉收缩减弱)。
- 反射中枢最易疲劳。
- 可能与递质的耗竭有关。
中枢抑制和中枢易化
突触后抑制
突触后抑制是指由中枢内抑制性中间神经元释放抑制性递质,通过产生 IPSP 对突触后神经元产生的抑制效应。

传入侧支性抑制(左)和回返性抑制(右)
- 传入侧支性抑制(交互性抑制):
- 联系方式:其神经联系方式是感觉传入纤维进入中枢后,在与反射通路上的某一中枢神经元形成兴奋性突触的同时,通过侧支与一个抑制性中间神经元也形成兴奋性突触(换元),这个抑制性中间神经元再与另一个中枢神经元形成抑制性突触。
- 举例:伸肌肌梭的传入冲动对与该肌相拮抗的屈肌运动神经元的抑制
- 意义:保证伸肌和屈肌活动的协调控制
- 回返性抑制:指神经元通过轴突侧支和抑制性中间神经元对自身的抑制。
- 联系方式:其神经联系方式是神经元兴奋时,传出纤维与,某一中枢神经元形成兴奋性突触的同时,通过轴突侧支兴奋一个抑制性中间神经元元,后者释放抑制性递质,反过来抑制原先发生兴奋的神经元及同一中枢的其他神经元。
- 举例:脊髓前角运动神经元的轴突支配骨骼肌,同时通过其轴突侧支与闰绍细胞构成突触联系,闰绍细胞再通过其短轴突回返性地抑制该运动神经元和同类的其他运动神经元
- 意义:及时终止神经元的活动,并使同一中枢内许多神经元的活动同步化
突触前抑制
一个神经元的轴突末梢与第二个兴奋性神经元的轴突末梢形成轴突-轴突式突触,前者兴奋时释放的递质就可影响后者兴奋时在其突触后的第三个神经元的胞体产生的兴奋性突触后电位。在这种突触联系模式中,若第一个神经元兴奋时释放的递质相对地降低了第二个神经元兴奋时(递质释放减少)在第三个神经元的胞体产生的兴奋性突触后电位,就称为突触前抑制。
- 突触前抑制可使最终的神经元(第三个神经元)产生的 EPSP 将明显减小(不会产生 IPSP)。
- 机制:
- 可能是第一个神经元末梢(A)兴奋时,释放 GABA 作用于第二个神经元末梢(B)上的 GABAA 受体,引起末梢 B 的 Cl- 电导增加(Cl- 外流),膜发生去极化,使传到末梢 B 的动作电位幅度变小(因为膜两侧电位差减小),时程缩短,结果使进入末梢 B 的 Ca2+ 减少,由此而引起递质释放量减少,最终导致第三个神经元的 EPSP 减小。
- 也可能是增加 K+ 通透性,复极化加快,减少 Ca2+ 内流
- 也可能是促代谢受体直接抑制递质释放
- 分布:广泛存在于中枢,尤其在感觉传入通路中,对调节感觉传入活动具有重要意义。
- 举例:脊神经后根感觉传入通路中存在突触前抑制
突触前易化
- 突触前易化与突触前抑制具有相同的结构基础
- 机制:可能是是轴突-轴突式突触的突舶前末梢释放某种递质(如 5-羟色胺),使末梢 A 内 cAMP 水平升高,钾通道发生磷酸化而关闭,结果导致动作电位的复极化过程延缓,Ca2+ 因动作电位时程延长而内流增多,使递质释放增加。
- 举例:突触性中的敏感化的发生机制即为突触前易化。
突触后易化
突触后易化表现为 EPSP 的总和,使 EPSP 幅度增大而更接近于阙电位水平,如果在此基础上给予一个刺激,就更容易达到阙电位水平而爆发动作电位。