声波通过外耳和中耳的传递到达耳蜗,经耳蜗的感音换能作用,最终将声波的机械能转变为听神经纤维上的神经冲动,后者上传到大脑皮层的听觉中枢,产生听觉。
- 听阈
- 在某一频率的声波,人耳刚能引起听觉的最小强度。
- 最大可听阈
- 在听阈以上继续增加强度,听觉的感受也相应增强,当强度增加到某一限度时,将引起鼓膜的疼痛感觉,这一限度称为最大可听阈。
- 听域
- 在每个频率的听阈和最大可听阈点上画曲线,两条曲线围成的面积,即为听域。
人耳能够感受的声波频率范围是 20–20000Hz,耳最敏感的声波频率范围为 1000–3000Hz,人的语言频率主要分布在 300–3000Hz 范围内。
外耳和中耳的功能
外耳的功能
- 耳郭:具有集音作用。
- 外耳道:
- 是声波传导的通道;
- 通过共振,使声压增强。
中耳的功能
- 组成:中耳由鼓膜、听骨链、鼓室和咽鼓管等结构组成。
- 鼓膜:呈顶点朝向中耳的浅漏斗状
- 是一个压力承受装置,其本身没有固有振动,但具有较好频率响应和较小失真度的特性,其振动与声波振动同始同终,几乎没有残余振动
- 听骨链:由锤骨、砧骨及镫骨依次连接而成
- 鼓膜:呈顶点朝向中耳的浅漏斗状
- 功能:中耳的主要功能是将声波振动能量高效地传给内耳,其中鼓膜和听骨链在声音传递过程中还起增压作用。
- 鼓膜和听骨链-增压作用:由于鼓膜和卵圆窗膜的面积比(17.2:1)和听骨链杠杆作用(长臂比短臂为 1.3:1),总共使声波在整个中耳传递过程中将增压 22.4 倍(17.2x1.3),而振幅约减小 1/4 。
- 若固定听骨链,中耳的增压作用将消失,声波在从空气传到内淋巴的过程中,约有 99.9% 被反射回空气,造成巨大的声能损失。
- 咽鼓管:为连接鼓室和鼻咽部的管道,其鼻咽部开口常处于闭合状态,当吞咽、打哈欠时开放,空气经咽鼓管进入鼓室,使鼓室内气压与外界大气压相同,以维持鼓膜的正常位置与功能(平衡鼓膜两侧压力)。
- 鼓膜张肌和镫骨肌:当声压过大时(70dB 以上),可反射性引起收缩,使鼓膜紧张,各听小骨之间的连接更为紧密,导致中耳传音效能降低,阻止较强的振动传到耳蜗,从而对内耳的感音装置起到保护作用。
- 由于完成上述反射需要 40–160 毫秒,故对突发性爆炸声的保护作用不大
- 鼓膜和听骨链-增压作用:由于鼓膜和卵圆窗膜的面积比(17.2:1)和听骨链杠杆作用(长臂比短臂为 1.3:1),总共使声波在整个中耳传递过程中将增压 22.4 倍(17.2x1.3),而振幅约减小 1/4 。
声波传入内耳的途径
- 气传导:
- 声波 → 鼓膜 → 听骨链 → 卵圆窗膜 → 耳蜗
- 声波 → 鼓膜 → 鼓室空气 → 圆窗膜 → 耳蜗
- 骨传导:声波直接作用于颅骨,经颅骨和耳蜗骨壁传入耳蜗。
异常状态下的传导
- 当鼓膜或中耳病变引起传音性耳聋时,气传导明显受损,而骨传导却不受影响,甚至相对增强
- 当耳蜗病变引起感音性耳聋时,音叉试验的结果表现为气传导和骨传导均异常
内耳耳蜗的功能
内耳又称迷路,位于颞骨岩部的骨质内,分为骨迷路和膜迷路。
- 骨迷路为骨性隧道,膜迷路为膜性结构,套在骨迷路内,形状与之相似。
- 骨迷路与膜迷路之间充满外淋巴,膜迷路内充满内淋巴,内、外淋巴互不相通。
迷路在功能上可分为耳蜗和前庭器官两部分。耳蜗的功能是将传到耳蜗的机械振动转变为听神经纤维的神经冲动。
耳蜗的功能结构要点
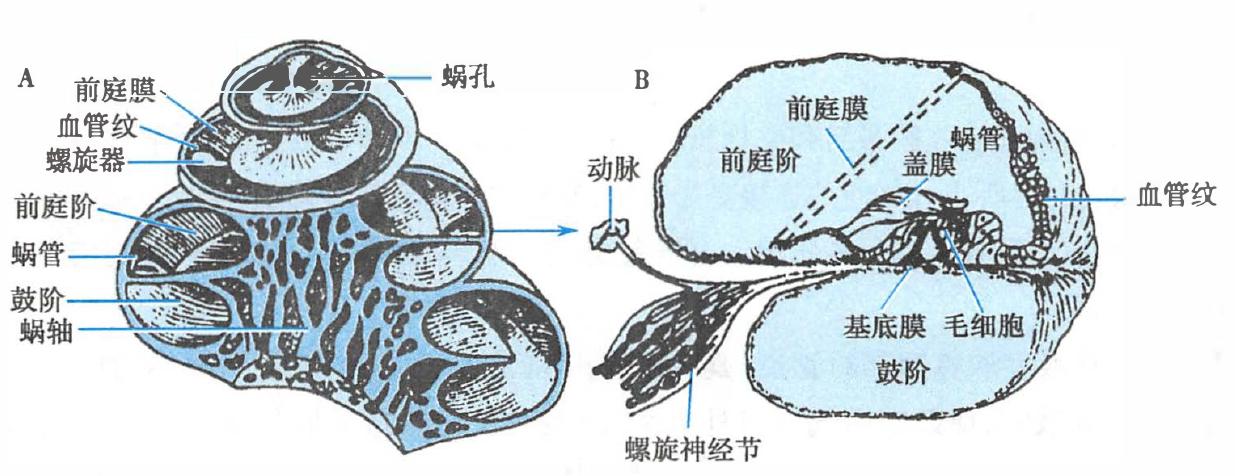
耳蜗纵切面(A)和耳蜗管横切面(B)示意图
耳蜗管被前庭膜和基底膜分成三个管腔,上方为前庭阶,中间为蜗管(也称中阶),下方为鼓阶。
- 前庭阶:在耳蜗底部与卵圆窗膜相接
- 蜗管(中阶):螺旋形的膜性盲管
- 鼓阶:在耳蜗底部与圆窗膜相接
内淋巴和外淋巴
- 蜗管是膜迷路的一部分,其内充满内淋巴;
- 前庭阶和鼓阶内都充满外淋巴,它们在蜗顶部通过蜗孔相沟通。
- 内淋巴与外淋巴不相通。
听觉感受器–螺旋器/柯蒂器
在基底膜上有听觉感受器,后者称为螺旋器或柯蒂器。
- 螺旋器由内、外毛细胞及支持细胞等组成:
- 毛细胞:
- 顶部与蜗管内淋巴接触
- 底部与鼓阶外淋巴相接触
- 底部与来自螺旋神经节的双极神经元周围突形成突触,而双极神经元中枢突则穿出蜗轴形成听神经
- 毛细胞:
- 其上覆以盖膜:盖膜在内侧与蜗轴相连,外侧则游离于内淋巴中。
耳蜗的感音换能作用
基底膜的振动和行波理论
不同频率的声波引起的行波都是从基底膜的底部开始,但声波频率不同,行波传播的距离和最大振幅出现的部位有所不同:声波频率越高,行波传播越近,最大振幅出现的部位越靠近蜗底;低频声波则反之。因此耳蜗底部受损时主要影响高频听力,而耳蜗顶部受损时则主要影响低频听力。
耳蜗的感音换能机制
感受器电位的产生
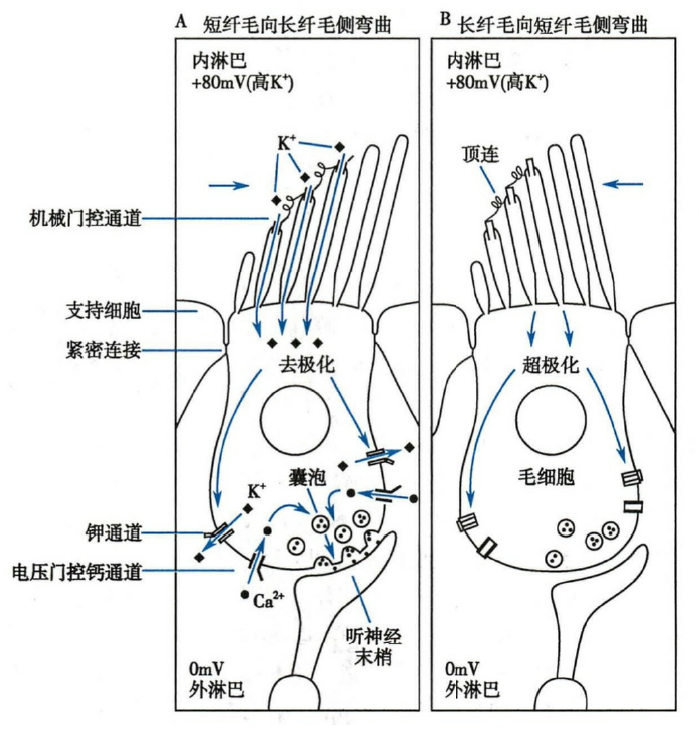
机械门控通道在毛细胞信号转导中的作用
当声波刺激引起基底膜振动时,盖膜与基底膜便沿着各自的轴上、下移动,于是在盖膜和基底膜之间产生剪切运动。
- 当基底膜上移时,短纤毛向长纤毛侧弯曲,引起通道开放,大量 K+ 内流(内淋巴高钾),由此产生去极化感受器电位;
- 而当基底膜下移时,长纤毛向短纤毛侧弯曲,引起通道关闭,K+ 内流终止而产生超极化感受器电位。
发生器电位及动作电位的产生
- 内毛细胞(也包括前庭器官中的毛细胞):
- 听神经传入纤维,90%–95% 分布到内毛细胞
- 当产生去极化感受器电位后,细胞基底侧膜上的电压门控钙通道被激活开放,引起 Ca2+内流,使细胞内 Ca2+浓度升高,触发递质释放,进而引起听神经纤维产生动作电位,并向听觉中枢传递
- 将不同频率的声波振动转变为听神经纤维动作电位,向中枢传送听觉信息
- 外毛细胞
- 听神经传入纤维,仅有 5%–10% 分布到外毛细胞
- 发生去极化时出现胞体缩短,发生超极化时则出现胞体伸长,外毛细胞的这种电-机械换能特性称为电能动性,是由膜上的马达蛋白 prestin 所驱动。Prestin 能感受细胞膜电位的变化,继而发生构象改变,导致外毛细胞缩短或伸长,从而增强基底膜的上移或下移
- 起到耳蜗放大器作用,可感知并迅速加强基底膜的振动,从而有助于盖膜下内淋巴的流动,使内毛细胞更易受到刺激,提高了对相应振动频率的敏感性
- prestin 失活,则外毛细胞失去耳蜗放大器作用,可引起动物耳聋
耳蜗的生物电现象
耳蜗内电位
- 当耳蜗未受刺激时,如果以鼓阶外淋巴的电位为参考零电位,则可测得蜗管内淋巴的电位为 +80 mV 左右,这一电位称为耳蜗内电位。此时毛细胞的静息电位为-70–-80mV。
- 外淋巴中含有较高浓度的 Na+ 和较低浓度的 K+,而内淋巴则正好相反,类似细胞内液和细胞外液。
- 毛细胞顶部浸浴在内淋巴(+80 mV,耳蜗内电位)中,而周围和底部则浸浴在外淋巴(0 mV,参考电位)中,毛细胞的静息电位为 -70–-80 mV,因此毛细胞顶端膜内、外的电位差可达 150–160 mV,而毛细胞周围和基底膜内、外的电位差仅约 80 mV。
- 内淋巴中正电位的产生和维持与蜗管外侧壁血管纹的活动密切相关。血管纹由边缘细胞、中间细胞和基底细胞所构成。
- 耳蜗内电位对基底膜的机械位移很敏感:
- 当基底膜向鼓阶方向位移时,耳蜗内电位可增高 10–15mV;
- 向前庭阶方向位移时,耳蜗内电位可降低 10 mV 左右。
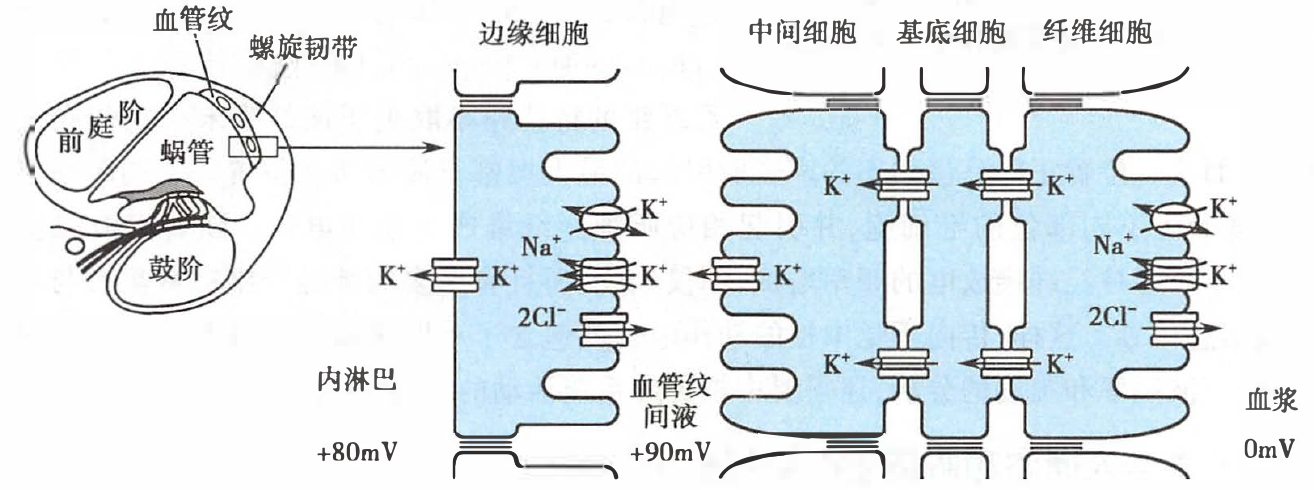
血管纹在产生和维持耳蜗内淋巴高钾中的作用机制示意图
- 螺旋韧带中的纤维细胞通过钠泵和 NKCC 同向转运体将 K+ 转入细胞内;
- 然后通过纤维细胞、基底细胞以及中间细胞三种细胞之间的缝隙连接,将 K+ 转入中间细胞内,使中间细胞内 K+ 浓度增高;
- 经中间细胞膜上的钾通道,将 K+ 转运到血管纹间液;
- 边缘细胞通过钠泵和 NKCC 同向转运体,将血管纹间液中的 K+ 转运到边缘细胞内,再经边缘细胞膜上的钾通道,将 K+ 转入内淋巴中。
临床上常用的依他尼酸和呋塞米等利尿药可通过抑制 NKCC 同向转运体,使内淋巴正电位不能维持,导致听力障碍。
耳蜗微音器电位
当耳蜗受到声音刺激时,在耳蜗及其附近结构可记录到一种与声波的频率和幅度完全一致的电位变化,称为耳蜗微音器电位。耳蜗微电位是多个毛细胞在接受声音刺激时所产生的感受器电位的复合表现,其特点有:
- 呈等级式反应,即其电位随着刺激强度的增加而增大。
- 耳蜗微音器电位无真正的阈值,没有潜伏期和不应期,不易疲劳,不发生适应现象。
- 在低频范围内,耳蜗微音器电位的振幅与声压呈线性关系,当声压超过一定范围时则产生非线性失真。
- 耳蜗微音器电位具有一定的位相性,即当声音的位相倒转时,耳蜗微音器电位的位相也发生倒转,而听神经干动作电位则不能。