RNA 聚合酶
真核生物有三种不同的 RNA 聚合酶,分别使用不同类型的启动子,并且需要不同类型的转录因子配合。
真核生物的三种 RNA 聚合酶
| RNA pol | 分布 | 合成 | 对 α-鹅膏蕈碱 |
|---|---|---|---|
| Ⅰ | 核仁 | (18S、28S、5.8S)rRNA 的前体——45S rRNA | 不敏感 |
| Ⅱ(最活跃) | 核内 | 前体 mRNA(hnRNA) 调控性非编码 RNA:lncRNA、miRNA 和 piRNA | 十分敏感 |
| Ⅲ | 核内 | tRNA,5S rRNA,核小 RNA(snRNA,属于核酶) | 比较敏感 |
转录过程
转录起始
顺式作用元件
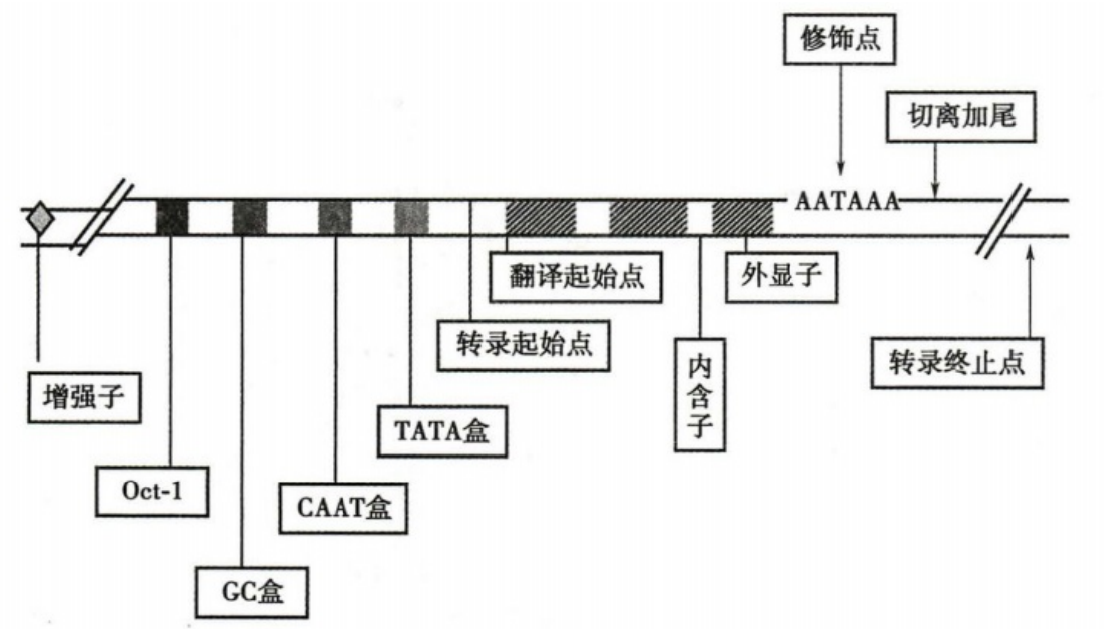
真核 RNA pol Ⅱ 识别的部分启动子共有序列
- 顺式作用元件
- 转录起始点上游的 DNA 序列,包括核心启动子序列、启动子上游元件、增强子。
- 核心启动子序列
- 转录起始点至上游-37bp的启动子区域
- 是转录起始前复合物的结合位点
- 起始点上游多数有共同的 TATA 序列,称为 Hognest 盒或 TATA 盒,通常认为这就是启动子的核心序列
- 启动子上游元件:
- 位于 TATA 盒上游的 DNA 序列,多在转录起始点上游约 40–200bp 的位置
- 比较常见的是位于-70bP 至-200bp 的 CAAT 盒和 GC 盒
- SP1 结合到 GC 盒上
- C/EBP 结合到 CAAT 盒上
- 这些元件与相应的蛋白因子结合能提高或改变转录效率
- 增强子
- 是能够结合特异基因调节蛋白并促进邻近或远隔特定基因表达的DNA序列
- 增强子距转录起始点的距离变化很大,但一般作用于最近的启动子
- 在所控基因的上游和下游都可发挥调控作用,但以上游为主
转录因子反式作用因子
- 反式作用因子
- 能直接、间接辨认和结合转录上游区段 DNA 或增强子的蛋白质,统称为反式作用因子,包括通用转录因子(基本转录因子,在真核生物进化中高度保守)和特异转录因子。
通用转录因子
通用转录因子指直接或间接结合 RNA pol 的一类转录调控因子。所有的 RNA pol Ⅱ 都需要通用转录因子,在真核生物进化中高度保守。
按“谈判桌模型”记忆- 结合 DNA:一个结合两个辅助
- TFⅡD:
- 含 TBP 亚基,结合启动子的 TATA 盒 DNA 序列
- 不是一种单一蛋白质,实际上是由 TATA 盒结合蛋白质(TBP)和 8–10 个 TBP 相关因子(TAF)组成的复合物
- TBP 支持基础转录但是不是诱导等所致的增强转录所必需的
- TAFs 对诱导引起的增强转录是必要的
- TFⅡB:
- 结合 TF Ⅱ D,稳定 TFⅡD-DNA 复合物
- 介导 RNA pol Ⅱ 的募集
- TFⅡA:
- 辅助和加强 TBP 与 DNA 的结合(非必需)
- TFⅡD:
- 酶活性:解激隔募调稳
- TFⅡH:
- 解旋酶(打开双链)和 ATPase 酶活性
- 作为蛋白激酶参与 CTD(羧基末端结构域)磷酸化,激活 RNA pol Ⅱ
- TFⅡF:
- 结合 RNA pol Ⅱ 并随其进入转录延长阶段,防止其与 DNA 的接触
- TFⅡE:
- 募集 TFⅡH 并调节其激酶和解螺旋酶活性
- 结合单链 DNA,稳定解链状态
- TFⅡH:
上游因子
- 定义:还与启动子上游元件(如GC盒、CAAT盒等顺式作用元件)结合的转录因子,称为上游因子
- 举例:如SP1结合到GC盒上,C/EBP结合到CAAT盒上
- 作用:调节通用转录因子与 TATA 盒的结合、RNA pol 在启动子的定位及起始复合物的形成,从而协助调节基因的转录效率
特异转录因子
- 是在特定类型的细胞中高表达,并对一些基因(奢侈基因)的转录进行时间和空间特异性调控的转录因子。
- 与远隔调控序列(如增强子等)结合的转录因子(反式作用因子)是主要的特异转录因子。
RNA polII 与启动子结合后启动转录,这需要多种蛋白质因子的协同作用
- 可诱导因子或上游因子与增强子或启动子上游元件的结合
- 辅激活因子和(或)中介子在可诱导因子、上游因子与通用转录因子 RNA polII 复合物之间起中介和桥梁作用
- 通用转录因子和 RNApolII 在启动子处组装成转录起始前复合物
转录起始前复合物
TFIID 的 TBP 亚基结合 TATA,在 TFIIA 和 IIB 的促进和配合下,形成 IID-IIA-IIB-DNA 复合体。
- 形成过程
- 先由TBP结合启动子的TATA盒,这时DNA发生弯曲,然后TFIIB与TBP结合,TFIIB也能与TATA盒上游邻近的DNA结合
- TFIIA不是必需的,其存在时能稳定已与DNA结合的TFIID-TBP复合体,并且在TBP与不具有特征序列的启动子结合时发挥重要作用
- TFIIB可以结合RNApolII
- TFIIB-TBP复合体再与由RNApolII和TFIIF组成的复合体结合
- TFIIF 的作用是通过和 RNApolII 一起与 TFIIB 相互作用,降低 RNApolII 与 DNA 的非特异部位的结合,来协助 RNApolII 靶向结合启动子
- 最后是TFIIE和TFIIH加入,形成闭合复合体,装配完成,这就是转录起始前复合物
- TFIIH
- 具有解旋酶活性,能使转录起始点附近的DNA双螺旋解开,使闭合复合体成为开放复合体,启动转录
- 还具有激酶活性,它的一个亚基能使 RNA pol Ⅱ 的 CTD 磷酸化
- CTD(羧基末端结构域)磷酸化
- 周期蛋白依赖性激酶 9(CDK9):是正性转录延长因子复合体的组成部分,对 RNA pol Ⅱ 的活性起正性调节作用
- CTD 磷酸化能使开放复合体的构象发生改变,启动转录
- 在转录延长期也很重要,而且影响转录后加工过程中转录复合体和参与加工附酵之间的相互作用
- 启动转录后,TFIID、TFIIA 和 TFIIB 等就会脱离转录起始前复合物。当合成一段含有 30 个左右核苷酸的 RNA 时,TFIIE 和 TFIIH 释放,RNApolII 进入转录延长期
- 在延长阶段,TFIIF 仍然结合 RNApolII,防止其与 DNA(非特异部位)的结合
少数几个反式作用因子的搭配启动特定的基因转录
数个反式作用因子(主要是可诱导因子和上游因子等转录因子)之间互相作用,再与基本转录因子、RNApol搭配而有针对性地结合、转录相应的基因
转录延长
- 因有核膜相隔,没有转录与翻译同步的现象
- 真核生物基因组 DNA 在双螺旋结构的基础上,与多种组蛋白组成核小体高级结构
- 转录延长可以观察到核小体移位和解聚现象
转录终止
- 真核生物的转录终止和转录后修饰密切相关
- 在可读框的下游,常有一组共同序列 AATAAA 序列,再下游还有相当多的 GT 序列,这些序列称为转录终止的修饰点
- 转录越过修饰点后,前体 mRNA 在修饰点处被核酸内切酶切断,随即加入 poly(A)尾及 5′ 帽子结构。下游的 RNA 虽继续转录,但很快被 RNA 酶降解
- 修饰点以后的转录产物无帽子结构,很快被降解,因此帽子结构是保护 RNA 免受降解的
- RNA pol 缺乏具有校读功能的 3′→5′ 核酸外切酶活性,因此转录发生的错误率比复制发生的错误率高
- 时间顺序:加帽 → 转录终止 → 加尾 → 剪接修饰