注意脂肪酸、胆固醇和酮体的合成虽然都是在合成脂类物质,但代表的生理意义不同:
- 脂肪酸、胆固醇的合成主要发生在机体糖充足甚至富余的情况;
- 而酮体生成主要发生在机体缺糖的情况,为保证脑等重要器官的糖分供给,利用脂肪生成酮体功能以节约葡萄糖。
甘油三酯代谢
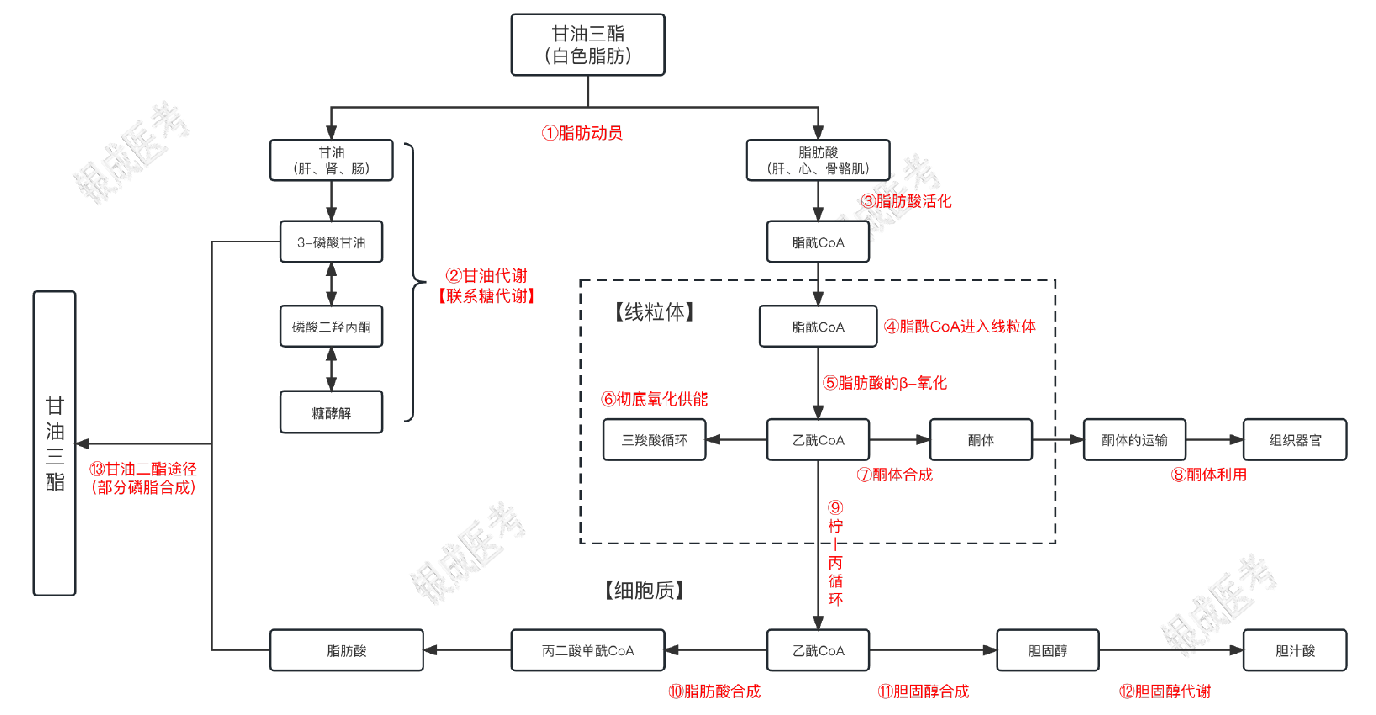
甘油三酯代谢汇总
多不饱和脂肪酸
多不饱和脂肪酸指含有两个及以上双键的脂肪酸,包括亚油酸、α-亚麻酸、花生四烯酸以及油酸和软油酸,其中前三种为必需脂肪酸。
- 提供必需脂肪酸:
- 必需脂肪酸:人体自身不能合成、必须由食物提供的脂肪酸(亚油酸、亚麻酸)。
- 花生四烯酸虽然能在人体内以亚油酸为原料合成,但消耗必需脂肪酸,一般也归为必需脂肪酸。
- 合成不饱和脂肪酸衍生物
- 前列腺素:
- PGE2 能诱发炎症,促进局部血管扩张,使毛细血管通透性增加,引起红、肿、痛、热等症状。
- PGE2,PGA2,能使动脉平滑肌舒张,有降血压作用。
- PGE2 及 PGI2 能抑制胃酸分泌,促进胃肠平滑肌蠕动。
- 卵泡产生的 PGE2,PGF2α,在排卵过程中起重要作用。PGF2α 可使卵巢平滑肌收缩,引起排卵。
- 子宫释放的 PGF2α 能使黄体溶解。
- 分娩时子宫内膜释放的 PGF2α 能使子宫收缩加强,促进分娩。
- 血管内皮细胞释放的 PGI2 有很强的舒血管以及抗血小板聚集作用
- 血栓烷 A2:
- 促进血小板聚集和血管收缩,促进凝血及血栓形成
- 白三烯:
- 过敏反应慢物质是 LTC4、LTD4 以及 LTE4 混合物,其支气管平滑肌收缩作用较组胺、PGF2α 强 100–1000 倍,作用缓慢而持久。
- 白三烯受体拮抗剂可以用来治疗哮喘的原因,既能缓解,也能控制,而且主要是控制效应
- LTB4 能调节白细胞功能,促进其游走和趋化作用,刺激腺苷酸环化酶,诱发多形核白细胞脱颗粒,使溶酶体释放水解酶类,促进炎症及过敏反应发展。
- IgE 与肥大细胞表面受体结合后,可引起肥大细胞释放 LTC4、LTD4 以及 LTE4,这三种物质能引起支气管及胃肠道平滑肌剧烈收缩,LTD4 还能使毛细血管通透性增加。
- 过敏反应慢物质是 LTC4、LTD4 以及 LTE4 混合物,其支气管平滑肌收缩作用较组胺、PGF2α 强 100–1000 倍,作用缓慢而持久。
- 前列腺素:
甘油三酯的分解
脂肪(甘油三酯)分解
脂肪动员
$$\ce{TG ->[ATGL] 脂肪酸 + DG ->[HSL] 脂肪酸 + MG ->[MGL] 脂肪酸 + 甘油}
\,.$$
- TG、DG、MG:甘油三酯、甘油二酯、甘油一酯
- ATGL:脂肪组织甘油三酯脂肪酶
- HSL:激素敏感甘油三酯脂肪酶(历史遗留问题)
- MGL:甘油一酯脂肪酶
- 禁食、饥饿或交感神经兴奋时,肾上腺素、去甲肾上腺素、胰高血糖素分泌增加
- 作用于白色脂肪细胞膜受体(G 蛋白偶联受体),激活腺苷酸环化酶(AC),使 cAMP 增多,激活 cAMP 依赖蛋白激酶
- 使细胞质内脂滴包被蛋白-1(Perilipin-1)和 HSL 磷酸化。
- 脂解激素:肾上腺素、去甲肾上腺素、胰高血糖素,能启动脂肪动员、促进脂肪分解
- 抗脂解激素:胰岛素、前列腺素 E2
甘油的利用
- 甘油可直接经血液运输至肝、肾、肠等组织利用,脂肪和骨骼肌的甘油激酶活性很低,对甘油的利用很有限。
- 甘油激酶的作用下变为 3-磷酸甘油,然后脱氢生成磷酸二羟丙酮,进入糖酵解,主要在肝脏利用
脂肪酸的利用
游离脂肪酸不溶于水,不能直接在血浆中运输。血浆清蛋白具有结合游离脂肪酸的能力(每分子清蛋白可结合 10 分子游离脂肪酸),能将脂肪酸运送至全身,主要由心、肝、骨骼肌等摄取利用。
脂肪酸活化
脂肪酸被氧化前必须先活化,由内质网、线粒体外膜上的脂酰 CoA 合成酶催化生成脂酰 CoA,消耗 1 个 ATP、2 个高能磷酸键,还需 CoA-SH 及 Mg2+ 参与。
脂酰 CoA 进入线粒体
脂酰 CoA 进入线粒体是脂肪酸 β-氧化的限速步骤,肉碱脂酰转移酶 Ⅰ 是脂肪酸 β-氧化的关键酶。
β-氧化:脂酰 CoA 分解产生乙酰 CoA 和还原当量
- 脱氢:脂酰 CoA 脱氢酶催化,生成 FADH2(1.5 ATP)
- 加水:烯酰 CoA 水化酶
- 脱氢:L-β-羟脂酰 CoA 脱氢酶,生成 NADH2(2.5 ATP)
- 硫解:β-酮硫解酶,加 CoASH 使碳链在β位断裂,生成 1 分子乙酰 CoA,和少 2 个碳原子的脂酰 CoA。
1 分子 2n C 的脂肪酸经历 n-1 次 β 氧化,生成 n 分子 乙酰 CoA,算上活化所需的 2 ATP,总共产生能量
$$Q_{2n} = (1.5 + 2.5)(n - 1) + 10n - 2 = 14n - 6(ATP)
\,.$$
如软脂酸(16 C)、硬脂酸(18 C)完全氧化分别产生 $14\times8-6 = 106$ 和 $14\times9 - 6 = 120$ 分子 ATP。
特殊脂肪酸的氧化方式
- 不饱和脂肪酸β-氧化需要转变构型:天然不饱和脂肪酸的双键为顺式,需要在异构酶的作用下变为反式才能继续氧化
- 超长碳链脂肪酸需要现在过氧化酶体氧化成比较短的碳链脂肪酸:这里氧化脱下来的 H 不是进行氧化磷酸化,而是与 O2 结合生成 H2O2
- 奇数碳原子的脂肪酸氧化会剩下丙酰 CoA:会转变成琥珀酰 CoA 进入三羧酸循环,可以进行糖异生
- 脂肪酸氧化还可以从远侧甲基端进行(ω-氧化)
酮体的合成与利用
酮体合成
酮体生成以脂肪酸 β-氧化生成的乙酰 CoA(葡萄糖、氨基酸代谢产生的乙酰 CoA 不会用于合成酮体)为原料,在肝脏线粒体由酮体合成酶系催化完成。
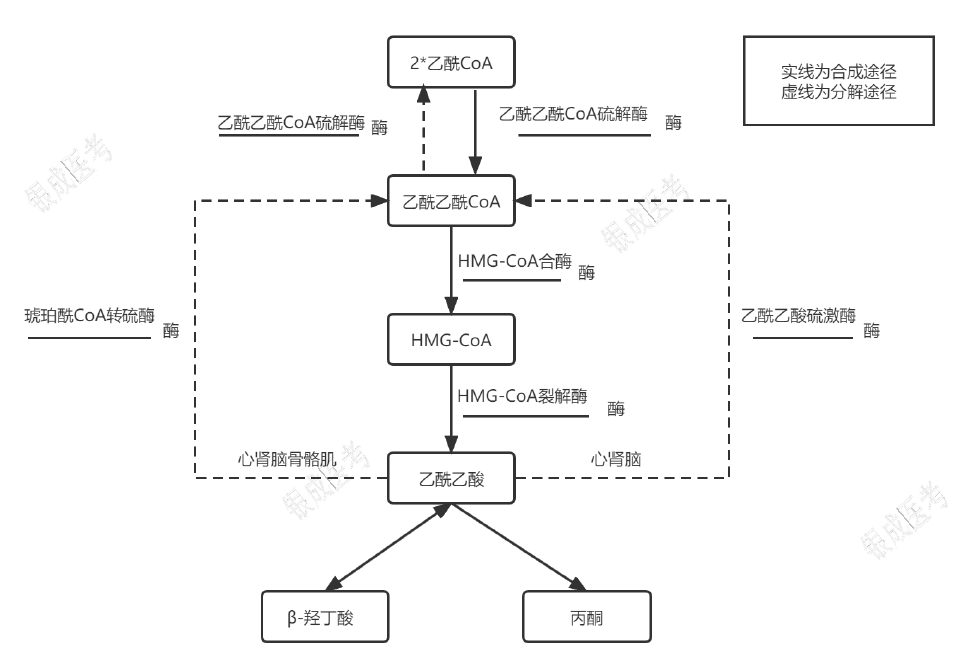
酮体合成
β-羟丁酸比乙酰乙酸多一个还原当量,是机体内酮体的主要组成(70%)和运输形式。
酮体的利用
- 给乙酰乙酸加巯基-CoA(-S-CoA)使其转变为乙酰乙酰 CoA
- 重要器官:心、肾(终于不是冤种了)、脑:乙酰乙酸硫激酶或琥珀酰 CoA 转硫酶
- 骨骼肌(骨骼肌只会抢):只能用琥珀酰 CoA 转硫酶
- 乙酰乙酰 CoA 硫解生成乙酰 CoA:由乙酰乙酰 CoA 硫解酶(酮体合成第一步的逆反应)催化,再给乙酰乙酰 CoA 加一个 -CoA-SH(由 CoA-SH 提供)使其裂解为 2 分子的乙酰 CoA。
酮体的意义
- 酮体是肝向外输出能量的重要形式
- 肝组织有活性较强的酮体合成酶系,但缺乏利用酮体的酶系。
- 心肌和肾皮质利用酮体能力大于利用葡萄糖的能力。
- 脑组织不能分解脂肪酸,却能有效利用酮体,在葡萄糖供应不足或利用障碍时,酮体是脑组织主要的能源物质。
- 饥饿或者糖尿病
- 酮症酸中毒:人体内血酮高出正常人数十倍,导致酸中毒。
- 酮尿:血酮体超过肾阈。
- “烂苹果气味”:血丙酮含量增大,通过呼吸道排出。
酮体生成的调节
- 餐饮状态:
- 饱餐后胰岛素分泌增加,脂解作用抑制,酮体生成减少
- 饥饿时,胰高血糖素等脂解激素分泌增多,酮体生成增多
- 糖代谢-:
- 糖供给充分时,酮体生成被抑制。
- 糖利用障碍或者饥饿时,脂肪酸分解加强,酮体增多。
- 丙二酸单酰 CoA 抑制酮体生成:说明糖代谢旺盛,在合成脂肪(酮体合成的原料来源于脂肪酸 β-氧化),故抑制酮体生成。
甘油三酯的合成
甘油三酯合成在细胞质中完成,以肝合成能力最强。但肝细胞不能储存甘油三酯(脂肪细胞是体内储存甘油三酯的“脂库”),需与载脂蛋白 B100、C 等载脂蛋白及磷脂、胆固醇组装成极低密度脂蛋白(VLDL),分泌入血,运输至肝外组织包。营养不良、中毒,以及必需脂肪酸、胆碱或蛋白质缺乏等可引起肝细胞 VLDL 生成障碍,导致甘油三酯在肝细胞蓄积,发生脂肪肝。
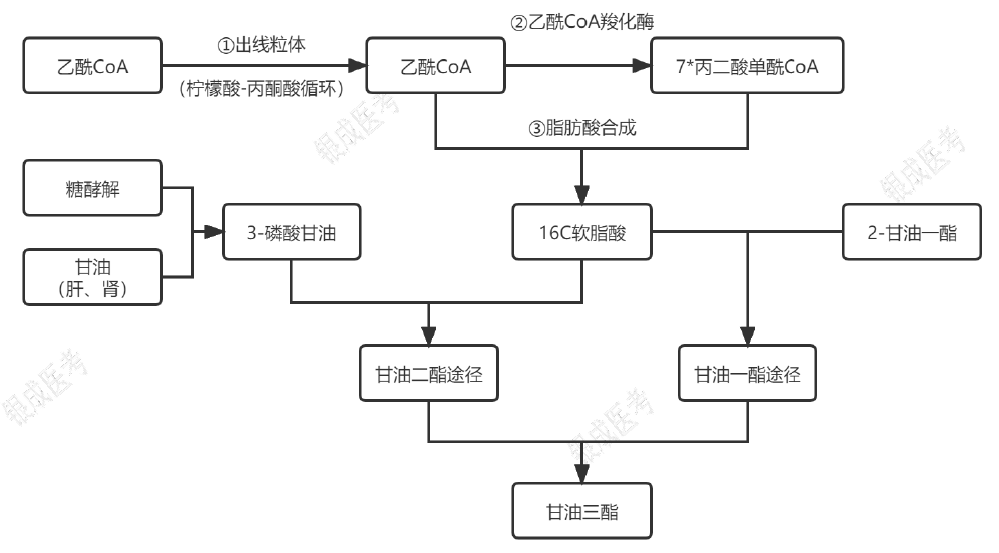
甘油三酯的合成
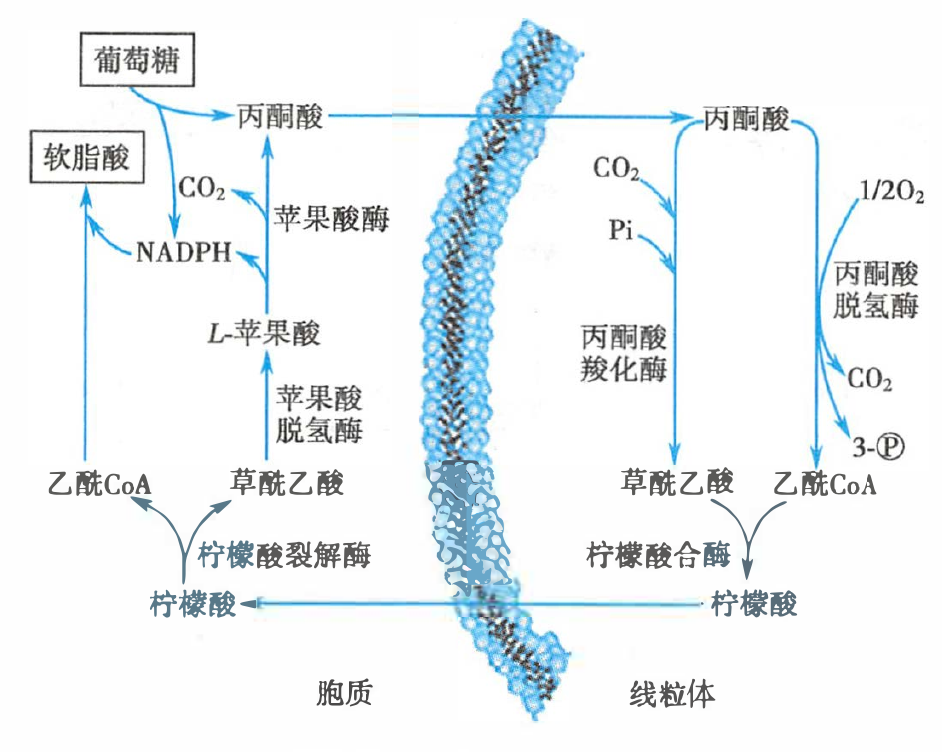
柠檬酸-丙酮酸循环示意图
柠檬酸-丙酮酸循环中的物质转换
甘油三酯合成的原料
- 3-磷酸甘油:
- 磷酸二羟丙酮:机体通过糖酵解途径,分解葡萄糖产生的磷酸二羟丙酮生成 3-磷酸甘油。
- 甘油:
- 肝、肾等组织含有甘油激酶,可催化游离甘油生成 3-磷酸甘油。
- 脂肪和骨骼肌的甘油激酶活性很低,不能利用甘油(脂肪动员)合成 3-磷酸甘油(甘油的利用)(
一边分解一边合成),因而不能利用甘油合成甘油三酯。
- 脂肪酸:
- 内源性:利用葡萄糖分解代谢中间产物乙酰 CoA 合成脂肪酸。
- 外源性:小肠黏膜细胞主要利用摄取的甘油三酯消化产物重新合成甘油三酯,当其以乳糜微粒形式运送至脂肪组织、肝等组织/器官后,脂肪酸亦可作为这些组织细胞合成甘油三酯的原料。
- 极低密度脂蛋白(VLDL):脂肪组织还可水解 VLDL 甘油三酯,释放脂肪酸用于合成甘油三酯。
甘油三酯合成的过程
- 软脂酸在细胞质中合成
- 乙酰 CoA 转化为丙二酸单酰 CoA(限速步骤)。
- 脂肪酸合成的关键酶:乙酰 CoA 羧化酶以 Mn2+ 为激活剂,含生物素辅基,消耗 1 分子 ATP
- 调节:
- 别构调节:
- 别构激活:柠檬酸、异柠檬酸(糖代谢活跃)
- 别构抑制:长链脂酰 CoA(脂肪分解活跃)
- 化学修饰
- 胰高血糖素、AMP[通过激活 AMP 敏感的蛋白激酶(AMPK)进行磷酸化修饰,注意不是常见的别构调节]可以通过磷酸化修饰的方式使其失活
- 胰岛素可以通过去磷酸化的方式使其恢复活性
- 高糖饮食:可以促进乙酰 CoA 羧化酶蛋白合成,增加酶的活性
- 别构调节:
- 由 1 分子乙酰 CoA 和 7 分子丙二酸单酰 CoA 合成 1 分子软脂酸
- 过程:缩合-还原-脱水-再还原(β-氧化:脱氢-加水-再脱氢-硫解),共消耗 14 分子 NADPH
- NADPH 来源:磷酸戊糖途径(葡糖-6-磷酸脱氢酶、6-磷酸葡萄糖酸脱氢酶)、柠檬酸-丙酮酸循环中的苹果酸酶(
苹果酸脱氢酶)
- 乙酰 CoA 转化为丙二酸单酰 CoA(限速步骤)。
- 软脂酸延长在内质网和线粒体进行
- 内质网:以丙二酸单酰 CoA 为原料,可延长至 24 碳
- 线粒体但是一般还是认为脂肪酸合成不在线粒体中进行:以乙酰 CoA 为原料,可延长至 24–26 碳,但以 18 碳的硬脂酸居多
- 甘油三酯的组装:
- 脂肪酸活化:在脂酰 CoA 合成酶的催化和 Mg2+ 的参与下,活化为脂酰 CoA,消耗 2 ATP(1 ATP,2 个高能磷酸键)
- 小肠黏膜细胞以甘油一酯途径合成甘油三酯:由脂酰 CoA 转移酶催化,ATP 供能,将脂酰 CoA 上的脂酰基转移至 2-甘油一酯羟基上合成甘油三酯。
- 肝和脂肪细胞以甘油二酯途径合成甘油三酯:也由脂酰 CoA 转移酶催化、ATP 供能,以磷酸二羟丙酮(糖酵解途径产生)还原生成的 3-磷酸甘油为起始物,合成 1,2-甘油二酯,再合成甘油三酯。见甘油磷脂合成。
胆固醇代谢
胆固醇合成
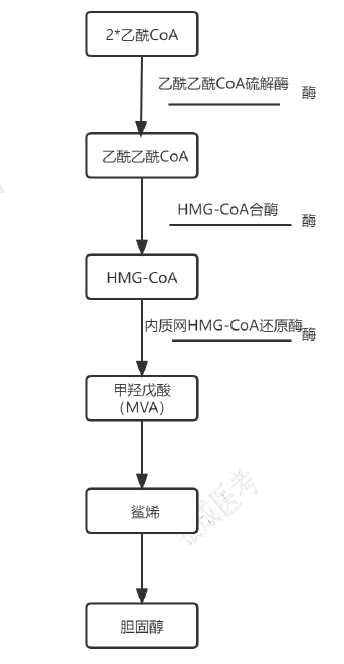
胆固醇合成
- 合成部位:光面内质网、细胞质
- 原料:以乙酰 CoA 为碳源,需要 NADPH 供氢、ATP 供能,合成 1 分子胆固醇需要 18 分子乙酰 CoA、36 分子 ATP 及 16 分子 NADPH。
胆固醇合成的调节
- 关键酶:HMG-CoA 还原酶
- 调节方式
- 产物抑制,底物促进;饥饿抑制,饱餐促进;能量少抑制,能量多促进
- 激素调节:
- 促进合成:胰岛素、甲状腺激素(也可胆汁酸的合成,综合作用使胆固醇含量降低)
- 抑制合成:胰高血糖素
甲状腺素的双相作用
胆固醇的去路
- 主要是转化为胆汁酸
- 还可以转化为类固醇激素:
- 肾上腺皮质激素:肾上腺皮质球状带、束状带及网状带细胞以胆固醇为原料分别合成醛固酮、皮质醇及雄激素。
- 性激素:睾丸间质细胞以胆固醇为原料合成睾酮等雄激素;卵泡内膜细胞及黄体以胆固醇为原料合成雌二醇及孕酮。
- 维生素 D3:胆固醇可在皮肤被氧化为 7-脱氢胆固醇,经紫外线照射转变为维生素 D3。
磷脂代谢
甘油磷脂
甘油磷脂合成
甘油磷脂合成过程
- 甘油、脂肪酸:来源于葡萄糖
- 多不饱和脂肪酸:是必需脂肪酸,只能从食物(植物油)中获取
- 胆碱:可由食物提供,或者由丝氨酸/甲硫氨酸合成
- 丝氨酸:可以脱羧生成乙醇胺
- 乙醇胺:可以从 SAM 获得 3 个甲基形成胆碱;可以羧化形成丝氨酸
- ATP:供能
- CTP:活化乙醇胺、胆碱、甘油二酯
甘油磷脂降解
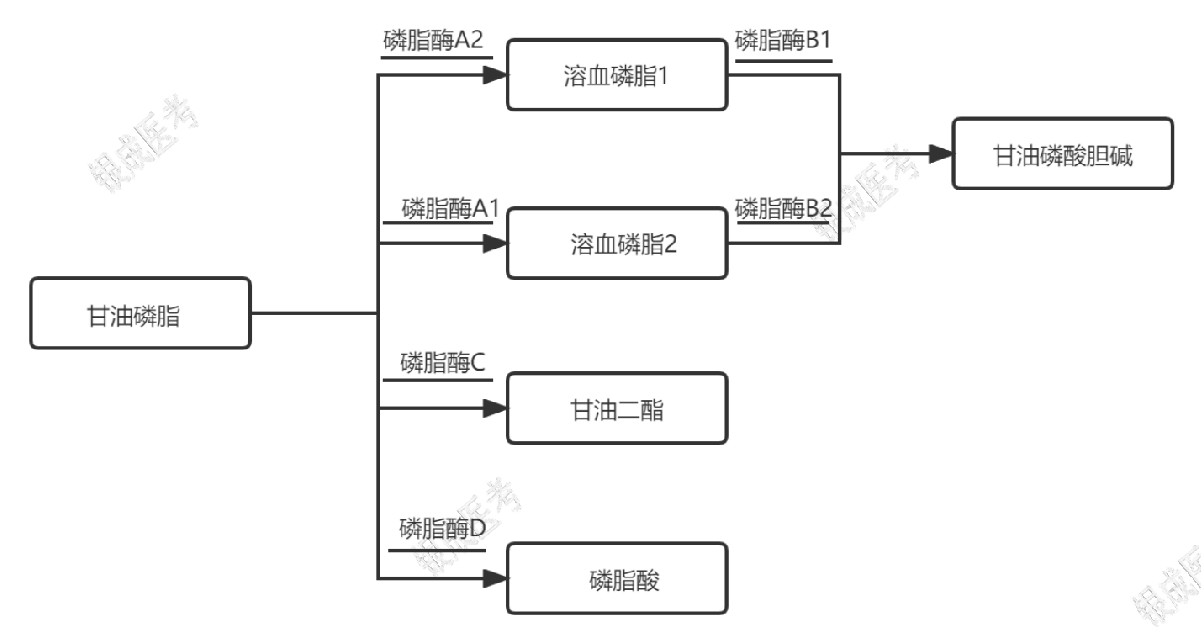
甘油磷脂的分解
记忆方法:1 不是 1,2 不是 2;Double 不是 D,aCid 不是酸。
鞘磷脂
鞘脂
- 鞘氨醇的氨基以酰胺键与 1 分子脂肪酸结合成神经酰胺,为鞘脂的母体结构。
- 鞘脂的结构通式如下,因取代基-X 不同,可分为两类:
- 鞘磷脂:取代基为磷酸胆碱、磷酸乙醇胺
- 鞘糖脂:取代基为葡萄糖、半乳糖、唾液酸等
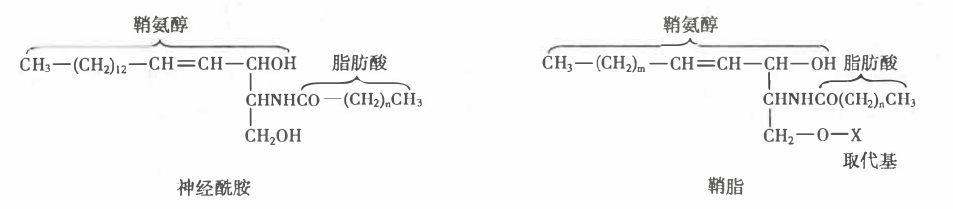
神经酰胺和鞘脂
含鞘氨醇或二氢鞘氨醇的磷脂称为鞘磷脂。
- 人体各组织细胞内质网均存在合成鞘氨醇酶系,以脑组织活性最高。
- 神经鞘磷脂是人体含量最多的鞘磷脂,由鞘氨醇(
甘油)、脂肪酸及磷酸胆碱构成。- 合成鞘氨醇的基本原料是软脂酰 CoA、丝氨酸和胆碱,还需磷酸吡哆醛、NADPH 及 FAD 等辅酶参加。
血浆脂蛋白代谢
血浆脂蛋白对比
| 乳糜微粒 | 极低密度脂蛋白 前 β-脂蛋白 | 低密度脂蛋白 β-脂蛋白 | 高密度脂蛋白 α-脂蛋白 | |
|---|---|---|---|---|
| 电泳位置 | 原点(点样位) | α2-球蛋白 | β-球蛋白 | α1-球蛋白 |
| 合成部位 | 小肠黏膜细胞 | 肝细胞、小肠黏膜细胞 | 血浆 | 肝、小肠 |
| 原料来源 | 食物脂肪 | 葡萄糖代谢中间产物、 内源性/外源性脂肪酸 | 由 VLDL 转变而来 | |
| 主要成分 | TG,少量胆固醇 | TG,少量胆固醇 | 胆固醇 | 胆固醇 |
| 主要功能 | 转运外源性 TG、胆固醇 | 转运内源性 TG | 转运内源性胆固醇 | 逆向转运胆固醇 |
- 电泳距离:α1 球蛋白 > 前 β 蛋白 > β 蛋白 > CM
- 超速离心(密度)/蛋白质含量/脂质含量的倒序:HDL > LDL > VLDL > CM
- 胆固醇含量:LDL > HDL > VLDL > CM
CM 在餐后合成半衰期仅 5–15 min,约可在血中存在 12 h。
VLDL 生成障碍可导致甘油三酯在脂肪细胞中堆积,导致脂肪肝,见甘油三酯代谢。
脂蛋白异常血症分型
| 分型 | 血浆脂蛋白升高 | 甘油三酯 | 胆固醇 |
|---|---|---|---|
| Ⅰ | CM | ↑↑↑ | ↑ |
| Ⅱa | LDL | ↑↑ | |
| Ⅱb | LDL、VLDL | ↑↑ | ↑↑ |
| Ⅲ | IDL(电泳宽 β 带) | ↑↑ | ↑↑ |
| Ⅳ | VLDL | ↑↑↑ | |
| Ⅴ | VLDL、CM | ↑↑↑ | ↑ |