胞质内受体
能与该型受体结合的信号分子有类固醇激素、甲状腺激素、视黄酸和维生素 D 等。
- 核受体:当激素进入细胞后,如果其受体是位于细胞核内,激素被运输到核内,与受体形成激素-受体复合物
- 胞质内受体:
- 如果受体是位于细胞质中,激素则在细胞质中结合受体,导致受体的构象变化,与热激蛋白分离,并暴露出受体的核内转移部位及 DNA 结合部位,激素-受体复合物向细胞核内转移,穿过核孔,迁移进入细胞核内,并结合于其靶基因邻近的激素反应元件上
- 结合于激素反应元件的激素-受体复合物再与位于启动子区域的基本转录因子及其他的特异转录调节分子作用,从而开放或关闭其靶基因,进而改变细胞的基因表达谱
- 不同的激素-受体复合物结合于不同的激素反应元件
离子通道型受体
- 结构特征
- 离子通道是由蛋白质寡聚体形成的孔道,其中部分单体具有配体结合部位
- 通道的开放或关闭直接受化学配体的控制,称为配体门控受体型离子通道,其配体主要为神经递质
- 离子通道受体的典型代表是N型乙酰胆碱受体,α亚基具有配体结合部位
- 离子通道受体信号转导的最终效应是细胞膜电位改变。这类受体引起的细胞应答主要是去极化与超极化
- 离子通道受体是通过将化学信号转变成为电信号而影响细胞功能的
- 举例
- 阳离子通道:乙酰胆碱、谷氨酸、5-羟色胺的受体
- 阴离子通道:甘氨酸、γ-氨基丁酸的受体
- 阳离子通道和阴离子通道的差异是由于构成亲水性通道的氨基酸组成不同,因而通道表面携带有不同电荷所致
G 蛋白耦联受体
- G 蛋白偶联受体(GPCR)
- G 蛋白偶联受体在结构上为单体蛋白,氨基端位于细胞膜外表面,羧基端在胞膜内侧,其肽链反复跨膜七次,因此又称为七次跨膜受体。
- 此类受体通过 G 蛋白向下游传递信号,因此称为 G 蛋白偶联受体。
传递模式
G 蛋白偶联受体介导的信号转导途径具有相同的基本模式:
- 细胞外信号分子结合受体,通过别构效应将其激活
- 受体激活 G 蛋白,G 蛋白在有活性和无活性状态之间连续转换,称为 G 蛋白循环
- 活化的 G 蛋白激活下游效应分子
- 不同的α亚基激活不同的效应分子,如 AC、PLC 等效应分子都是由不同的 G 蛋白所激活
- 有的α亚基可以激活 AC 称为αs,有的α亚基可以抑制 AC 称为αi
- G 蛋白的效应分子向下游传递信号的主要方式是催化产生小分子信使
- 小分子信使作用于相应的靶分子(主要是蛋白激酶),使之构象改变而激活
- 蛋白激酶通过磷酸化作用激活一些与代谢相关的酶、与基因表达相关的转录因子以及一些与细胞运动相关的蛋白质,从而产生各种细胞应答反应
具体路径
不同 G 蛋白偶联受体可通过不同途径传递信号:
- cAMP-PKA 途径
- 以靶细胞内cAMP浓度改变和PKA激活为主要特征
- 胰高血糖素、肾上腺素、促肾上腺皮质激素等可激活此途径
- PKA活化后,可使多种蛋白质底物的丝氨酸/苏氨酸残基发生磷酸化,改变其活性状态,底物分子包括一些糖代谢和脂代谢相关的酶类、离子通道和某些转录因子
- 调节代谢
- 激活糖原磷酸化酶b激酶、激素敏感脂肪酶、胆固醇酯酶,促进糖原、脂肪、胆固醇的分解代谢
- 抑制乙酰CoA羧化酶、糖原合酶, 抑制脂肪合成和糖原合成
- 调节基因表达
- 激活后进入细胞核的PKA可使cAMP反应元件结合蛋白(CREB)磷酸化
- 磷酸化的CREB结合于cAMP反应元件(CRE),并与CREB结合蛋白(CBP)结合
- 与CREB结合后的CBP作用于通用转录因子(包括TFRB),促进通用转录因子与启动子结合,激活基因的表达
- 调节细胞极性:通过磷酸化作用激活离子通道,调节细胞膜电位
- IP3/DAG-PKC 途径
- 促甲状腺素释放激素、去甲肾上腺素、抗利尿素与受体结合后所激活的G蛋白可激活PLC
- PLC 水解膜组分 PIP2,生成 DAG 和 IP3
- IP3 促进细胞钙库内的 Ca2+ 迅速释放,使细胞质内的 Ca2+ 浓度升高
- Ca2+ 与细胞质内的 PKC 结合并聚集至质膜
- 质膜上的 DAG、磷脂酰丝氨酸与 Ca2+共同作用于 PKC 的调节结构域,使 PKC 变构而暴露出活性中心
- Ca2+/钙调蛋白依赖的蛋白激酶途径
- G蛋白偶联受体至少可通过三种方式引起细胞内Ca2+浓度升高
- 某些G蛋白可以直接激活细胞质膜上的钙通道
- 或通过PKA激活细胞质膜的钙通道,促进Ca2+流入细胞质
- 或通过IP3促使细胞质钙库释放Ca2+
- 细胞质中的Ca2+浓度升高后,通过结合钙调蛋白传递信号
- Ca2+/CaM复合物的下游信号转导分子是一些蛋白激酶,它们的共同特点是可被Ca2+/CaM复合物激活,因而统称为钙调蛋白依赖性蛋白激酶
- 钙调蛋白依赖性激酶属于蛋白质丝氨酸/苏氨酸激酶
- 肌球蛋白轻链激酶、磷酸化酶激酶、钙调蛋白依赖性激酶
- 可激活各种效应蛋白质,可在收缩和运动、物质代谢、神经递质的合成、细胞分泌和分裂等多种生理过程中起作用
- G蛋白偶联受体至少可通过三种方式引起细胞内Ca2+浓度升高
酶耦联受体
- 酶耦联受体
- 酶偶联受体主要通过蛋白质修饰或相互作用传递信号。
- 酶偶联受体主要是生长因子和细胞因子的受体。此类受体介导的信号转导主要是调节蛋白质的功能和表达水平、调节细胞增殖和分化。
蛋白激酶偶联受体介导的信号转导途径具有相同的基本模式
- 胞外信号分子与受体结合,导致第一个蛋白激酶被激活
- 有的受体自身具有蛋白激酶活性,此步骤是激活受体胞内结构域的蛋白激酶活性
- 有些受体自身没有蛋白激酶活性,此步骤是受体通过蛋白质-蛋白质相互作用激活某种蛋白激酶
- 通过蛋白质-蛋白质相互作用或蛋白激酶的磷酸化修饰作用激活下游信号转导分子,从而传递信号,最终仍是激活一些特定的蛋白激酶
- 蛋白激酶通过磷酸化修饰激活代谢途径中的关键酶、转录调控因子等,影响代谢途径、基因表达、细胞运动、细胞增殖等
常见的信号转导途径
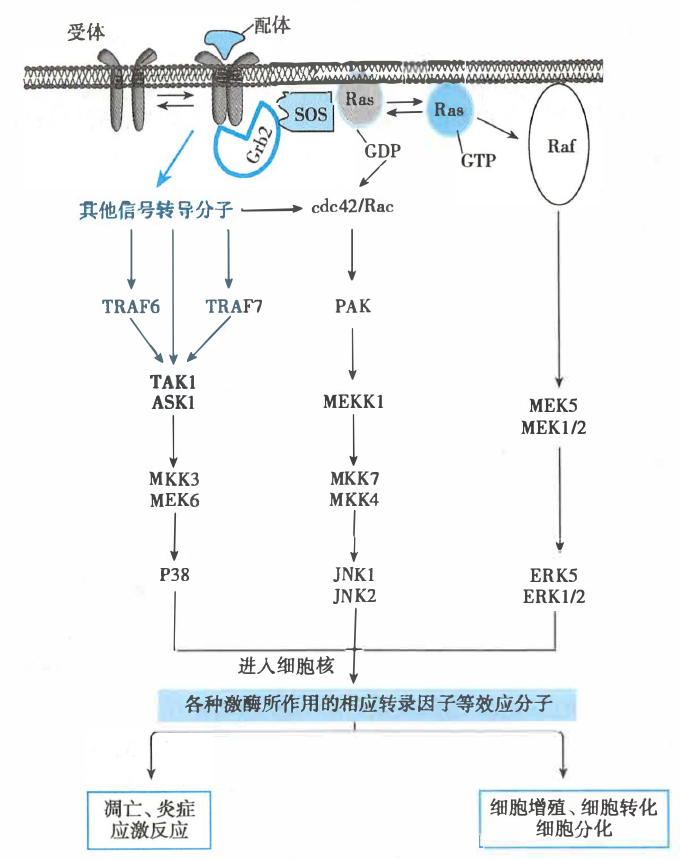
- Ras/MAPK 途径(胰岛素促生长):
- 受体与配体结合后形成二聚体,激活受体的蛋白激酶活性
- 受体自身酪氨酸残基磷酸化,形成 SH2 结合位点,从而能够结合含有 SH2 结构域的接头蛋白 Grb2
- Grb2 的两个 SH3 结构域与 SOS 分子中的富含脯氨酸序列结合,将 SOS 活化
- 活化的 SOS 结合 Ras 蛋白,促进 Ras 释放 GDP、结合 GTP
- 活化的 Ras 蛋白(Ras-GTP)可激活 MAPKKK(Raf),活化的 MAPKKK 可磷酸化 MAPKK(MEK)而将其激活,活化的 MAPKK 将 MAPK(ERK)磷酸化而激活
- 活化的 MAPK 可以转位至细胞核内,通过磷酸化作用激活多种效应蛋白,从而使细胞对外来信号产生生物学应答
- 其他途径:JAK-STAT 途径、Smad 途径、PI-3K 途径(胰岛素降糖)、NF-kB 途径等