原核生物基因组结构特点
- 基因组中很少有重复序列
- 编码蛋白质的结构基因为连续编码,且多为单拷贝基因,但编码 rRNA 的基因仍然是多拷贝基因
- 结构基因在基因组中所占的比例(约占 50%)远远大于真核基因组
- 许多结构基因在基因组中以操纵子为单位排列
操纵子是原核基因转录调控的基本单位
- 结构基因
- 通常包括数个功能上有关联的基因,它们串联排列,共同构成编码区
- 这些结构基因共用一个启动子和一个转录终止信号序列,因此转录合成时仅产生一条mRNA长链,为几种不同的蛋白质编码
- mRNA分子携带了几条多肽链的编码信息,被称为多顺反子mRNA
- 调控序列
- 启动子:
- 是 RNA 聚合酶结合的部位,是决定基因表达效率的关键元件
- 共有序列决定启动子的转录活性大小
- 操纵元件:
- 是一段能被特异的阻遏蛋白识别和结合的 DNA 序列
- 操纵序列(大概就是操纵元件)
- 是原核阻遏蛋白的结合位点
- 当操纵序列结合有阻遏蛋白时会阻碍 RNA 聚合酶与启动子的结合,或使 RNA 聚合酶不能沿 DNA 向前移动,阻遏转录,介导负性调节
- 原核操纵子调控序列中还有一种特异的 DNA 序列可结合激活蛋白,结合后 RNA 聚合酶活性增强,使转录激活,介导正性调节,如CAP 结合位点
- 启动子:
- 调节基因
- 编码能够与操纵元件结合的阻遏蛋白
- 阻遏蛋白可以识别、结合特异的操纵元件,抑制基因转录,所以阻遏蛋白介导负性调节
- 阻遏蛋白介导的负性调节机制在原核生物中普遍存在
- 还有一些调控蛋白质对原核基因转录调控起着重要的作用
- 特异因子
- 决定 RNA 聚合酶对一个或一套启动序列的特异识别和结合能力
- 激活蛋白
- 可结合启动子邻近的DNA序列,提高RNA聚合酶与启动序列的结合能力,从而增强RNA聚合酶的转录活性,是一种正性调节
- 分解(代谢)物基因激活蛋白(CAP)就是一种典型的激活蛋白
- 特异因子
乳糖操纵子是典型的诱导型调控
乳糖操纵子的结构
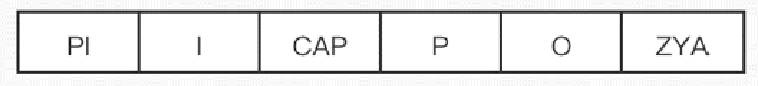
乳酸操纵子
- 结构基因:在环境中没有乳糖时,这些基因处于关闭状态,只有当环境中有乳糖时,这些基因才被诱导开放,合成代谢乳糖所需要的酶。
- Z:β-半乳糖苷酶
- Y:通透酶
- A:乙酰基转移酶
- 乳糖操纵子的调控区
- 操纵序列 O:结合阻遏蛋白,使操纵子受阻遏而处于关闭状态(负性调节启动子)
- 启动子 P
- 在启动子上游还有CAP结合位点(正性调节启动子)
- 调节基因 I
- I 基因具有独立的启动子(PI),编码一种阻遏蛋白
- 阻遏蛋白与 O 序列结合,使操纵子受阻遏而处于关闭状态
乳糖操纵子受到阻遏蛋白和CAP的双重调节
- 阻遏蛋白的负性调节:
- 别乳糖:
- 没有乳糖存在时:lac 操纵子处于阻遏状态
- I 序列在 PI 启动序列作用下表达的 Lac 阻遏蛋白与 O 序列结合,阻碍 RNA 聚合酶与 P 序列结合,抑制转录启动
- 有乳糖存在时:lac 操纵子可被诱导
- 真正的诱导剂并非乳糖本身
- 乳糖经通透酶催化、转运进入细胞,再经原先存在于细胞中的少数β-半乳糖苷酶催化,转变为别乳糖
- 别乳糖作为一种诱导剂分子,结合阻遏蛋白,使蛋白质构象变化,导致阻遏蛋白与 O 序列解离而发生转录
- 没有乳糖存在时:lac 操纵子处于阻遏状态
- 异丙基硫代半乳糖昔:
- 别乳糖的类似物,是一种作用极强的诱导剂,不被细菌代谢而十分稳定,因此在基因工程领域和分子生物学实验中被广泛应用。
- 别乳糖:
- CAP 的正性调节:
- 葡萄糖缺乏时:cAMP 浓度增高,cAMP 与 CAP 结合,这时 CAP 结合在 lac 启动序列附近的 CAP 位点,可刺激 RNA 聚合酶转录活性,使之提高 50 倍
- 有葡萄糖存在时:cAMP 浓度降低,cAMP 与 CAP 结合受阻,因此 lac 操纵子表达下降
- 协同调节:
- Lac 阻遏蛋白阻遏转录时,CAP 对该系统不能发挥作用
- 如果没有 CAP 存在来加强转录活性,即使阻遏蛋白从操纵序列上解离仍几无转录活性
色氨酸操纵子通过阻遏作用和衰减作用抑制基因表达
- 阻遏作用:粗略调节
- 细胞内无色氨酸时:阻遏蛋白不能与操纵序列结合,因此色氨酸操纵子处于开放状态,结构基因得以表达
- 细胞内色氨酸的浓度较高时:色氨酸作为辅阻遏物与阻遏蛋白形成复合物并结合到操纵序列上,关闭色氨酸操纵子,停止表达用于合成色氨酸的各种酶
- 转录衰减:精确调节
- 促进已经开始转录的 mRNA 合成终止
- 色氨酸操纵子还可通过转录衰减的方式抑制基因表达
- 这段前导序列称为【衰减子】
乳酸操纵子中结构基因的产物是利用乳酸的酶,而色氨酸操纵子中结构基因的产物则是色氨酸(利用色氨酸的酶)。
原核基因表达在翻译水平受到精细调控
- 蛋白质分子结合于启动子或启动子周围进行自我调节
- 调节蛋白结合mRNA靶位点,阻止核糖体识别翻译起始区,从而阻断翻译
- 自我控制:调节蛋白一般作用于自身mRNA,抑制自身的合成
- 翻译阻遏利用蛋白质与自身mRNA的结合实现对翻译起始的调控:调节蛋白结合到起始密码子上,阻断与核糖体的结合
- 反义RNA利用结合mRNA翻译起始部位的互补序列调节翻译起始:反义RNA含有与特定mRNA翻译起始部位互补的序列,通过与mRNA杂交,阻断30S小亚基对起始密码子的识别及与SD序列的结合,抑制翻译起始
- mRNA密码子的编码频率影响翻译速度:当基因中的密码子是常用密码子时,mRNA的翻译速度快,反之,mRNA的翻译速度慢