代谢的整体性
体内代谢过程互相联系形成整体
- 物质代谢与能量代谢相互关联
- 一般情况下供能以糖及脂肪为主
- 因疾病不能进食或无食物供给时,肝糖异生增强,蛋白质分解增强
- 如饥饿持续一周以上,长期糖异生增强使蛋白质大量分解,威胁生命
- 任一供能物质的分解代谢占优势,常能抑制其他供能物质的氧化分解
- 糖、脂蛋白质代谢通过中间代谢产物而互相联系
- 糖-脂:
- 葡萄糖可转变为脂肪酸:糖代谢生成乙酰 CoA,在胞质中用于脂肪酸合成
- 脂肪酸不能转变为葡萄糖:脂肪酸(偶数碳链)线粒体内 β-氧化生成的乙酰 CoA 不能生糖
- 甘油可以在肝、肾、肠等组织甘油激酶的作用下转变为磷酸甘油,进入糖酵解,但是量极少
- 糖供给不足或者糖代谢障碍时,生成大量酮体,而由于缺乏草酰乙酸导致大量酮体并不能进三羧酸循环氧化,在血中堆积,造成高酮血症
- 糖-氨基酸:糖与大部分氨基酸(除了生酮氨基酸)可以互相转变
- 氨基酸-脂
- 氨基酸都可以分解生成乙酰CoA,为脂质的合成提供原料
- 还可以作为磷脂合成的原料
- 但是脂肪酸、胆固醇不能变为氨基酸,仅能通过甘油异生为糖后转变为某些非必需氨基酸,但是量很少
- 氨基酸、磷酸戊糖是合成核苷酸的原料
- 嘌呤:甘氨酸、天冬氨酸、谷氨酰胺、一碳单位
- 嘧啶:天冬氨酸、谷氨酰胺、一碳单位
- 糖-脂:
“只进不出”的脂肪
糖、蛋白质(氨基酸)均可以转换为脂肪,但脂肪(甘油、奇数链脂肪氧化生成的丙酰 CoA 忽略不计)不能转换为糖或蛋白质(氨基酸)。
代谢的主要调节方式
调节关键酶的活性
关键酶活性决定整个代谢途径的速度和方向
- 快速调节:化学修饰、别构调节、酶原激活
- 迟缓调节:改变酶蛋白分子的合成/降解速度,改变细胞内酶的含量
主要物质代谢在人体内的分布
物质代谢 | 分布 | 物质代谢 | 分布 |
|---|---|---|---|
糖代谢 | 脂质代谢 | ||
| 有氧氧化、无氧氧化 | 组织偏好 | 脂肪动员 | 白色脂肪 |
| 磷酸戊糖途径 | 脂质合成旺盛的组织(肝、 脂肪、哺乳期乳腺),增 殖活跃的组织(骨髓、肿 瘤),红细胞 | 甘油利用 | 肝(甘油激酶) |
| 糖原合成和分解 | 肝、骨骼肌 | 脂肪分解 | 除脑外的大多数组织,以 肝、心肌、骨骼肌能力最强 |
| 糖异生 | 肝(主要)、肾 | 酮体生成 | 肝 |
氨基酸代谢 | 酮体利用 | 心、肾、脑、骨骼肌 | |
| 转氨基作用 | 肝(ALT)、心肌(AST) | 甘油三酯合成 | 甘油二酯途径:肝、脂肪组织 甘油一酯途径:小肠 |
| L-谷氨酸脱氢酶 | 肝、肾、脑等 | 甘油磷脂合成 | 各组织,以肝、肾、肠等活 性最高 |
| 尿素合成 | 肝(尿素合成特异性器官) | 鞘氨醇合成 | 各组织,以脑活性最高 |
核苷酸代谢 | 胆固醇合成 | 各组织,主要是肝 | |
| 嘌呤核苷酸合成 | 从头合成:肝(主要)、 小肠黏膜、胸腺 补救合成:脑、骨髓 | 胆固醇转化 | 肝;肾上腺皮质、睾丸、卵 巢;皮肤 |
| 嘧啶核苷酸合成 | 主要是肝 |
主要代谢途径(多酶体系)在细胞内的分布
| 分布 | 代谢途径(多酶体系) |
|---|---|
| 细胞核 | DNA、RNA 合成 |
| 细胞质 | 糖原合成,脂肪酸合成,糖酵解,磷酸戊糖途径 |
| 内质网 | 磷脂合成、@脂肪酸延长(≤ 24 碳) |
| 线粒体 | 三羧酸循环,氧化磷酸化、脂肪酸延长(24–26 碳) |
| 内质网 + 细胞质 | 蛋白质合成,胆固醇合成 |
| 细胞质 + 线粒体 | 血红素合成,尿素合成,糖异生,脂肪酸氧化 |
| 溶酶体 | 多种水解酶 |

- 别构调节通过别构效应改变关键酶活性:代谢产物的堆积表明其代谢过强,超过了需求,常可使其代谢途径的关键酶受到别构抑制,从而降低整个代谢途径的强度,避免产生超过需要的产物
- 化学修饰通过酶促共价修饰调节酶的活性:
- 磷酸化/去磷酸化、乙酰化/去乙酰化、化学修饰的方式、甲基化/去甲基化、腺苷化/去腺苷化、-SH 与-S-S-互变
- 磷酸化/去磷酸化最多见,通常需要消耗能量,酶蛋白分子中丝氨酸、苏氨酸以及酪氨酸 苏苏洛 的羟基是磷酸化修饰的位点。
- 酶促化学修饰由于具有酶促反应的放大效应,催化效率比变构调节高
激素通过特异性受体调节靶细胞代谢
通过神经-体液途径协调整体代谢
- 饱餐状态:
- 高糖饮食
- 胰岛素明显水平升高,葡萄糖转变为糖原、甘油三酯等物质进行存储
- 高蛋白饮食
- 胰岛素水平中度升高,胰高血糖素水平升高,共同作用使得肝糖原分解补充血糖、供应脑组织
- 小肠吸收的氨基酸在肝异生为糖
- 部分氨基酸转化为乙酰CoA合成甘油三酯
- 高脂饮食
- 胰高血糖素升高,胰岛素水平降低
- 肝糖原分解补充血糖
- 肌肉组织氨基酸分解、转化为丙酮酸,输送至肝异生为糖
- 高糖饮食
- 空腹状态:
- 餐后 12 小时后
- 胰岛素水平降低,胰高血糖素升高
- 餐后 6–8 小时
- 肝糖原已经开始在胰高血糖素的作用下分解血糖
- 餐后 12–18 小时
- 肝糖原即将耗尽,主要靠糖异生补充血糖,脂肪动员中度增加
- 餐后 12 小时后
- 饥饿状态:
- 短期饥饿(1-3天未进食)
- 糖氧化供能减少,脂肪动员加强
- 机体从葡萄糖氧化供能转变为脂肪氧化供能为主
- 肝糖异生明显增强,主要来源于氨基酸
- 骨骼肌蛋白质分解加强
- 长期饥饿(未进食3天以上,饥饿4-7天)
- 脂肪动员进一步加强(可引起酮症酸中毒等严重代谢紊乱而致死)
- 蛋白质分解减少
- 糖异生明显减少
- 短期饥饿(1-3天未进食)
- 应激状态:与糖皮质激素的生理功能一致
- 肝糖原分解加强,血糖升高;肌糖原分解加强,血乳酸增加
- 脂肪动员加强,甘油、游离脂肪酸增加,酮体生成增加
- (肌)蛋白质分解加强,血氨基酸浓度升高
人体主要器官代谢特点
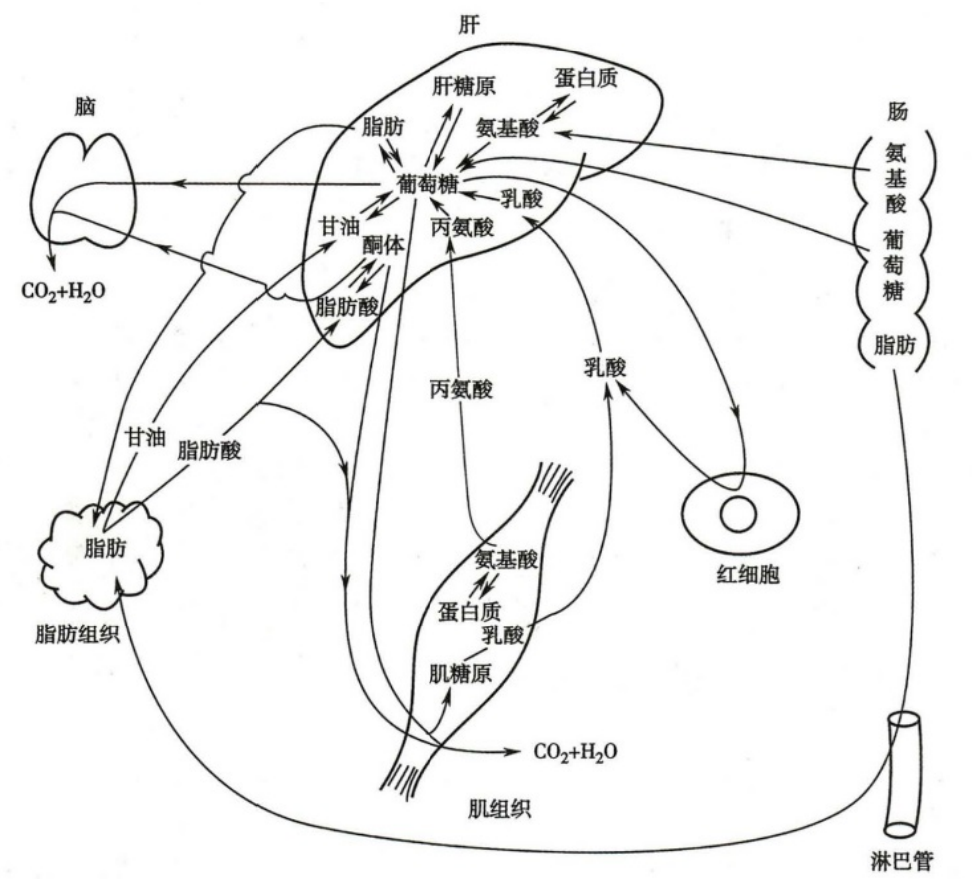
人体内主要器官代谢特点
- 肝
- 可以消耗葡萄糖、脂肪酸、甘油、氨基酸等,但不能利用酮体
- 肝合成的糖原可达肝重的5%
- 具有糖异生、酮体生成等特异性的代谢方式
- 可以大量合成脂肪,但不能存储脂肪,肝细胞合成的脂肪随合成的VLDL释放入血
- 脑
- 葡萄糖和酮体是主要能量物质
- 脑耗氧量高达全身总耗氧量1/4,是静息状态下单位重量组织耗氧量最大的器官
- 具有特异的氨基酸及其代谢调节机制
- 心肌
- 含有多种硫激酶,心肌优先利用脂肪酸分解供能
- 也有丰富的酮体利用酶,使用酮体供能
- 心肌富含LDH1,有利于乳酸氧化供能
- 所以会抑制葡萄糖酵解
- 所以主要通过有氧氧化,氧化脂肪酸,酮体和乳酸,极少进行糖酵解
- 在饱餐状态下,不排斥利用葡萄糖,餐后数小时或饥饿时用脂肪酸和酮体,运动后利用乳酸
- 骨骼肌
- 不同骨骼肌产能方式不同
- 红肌(持久):耗能多,主要靠氧化磷酸化供能
- 白肌(爆发):耗能少,主要靠糖酵解供能
- 骨骼肌适应不同耗能状态选择不同能源
- 直接能量来源:ATP(含量有限,不足以维持持续、剧烈运动)
- ATP消耗后:磷酸肌酸(间接利用)将高能磷酸键转移给ADP,生成ATP
- 储备:肌糖原(不能补充血糖)
- 静息状态下:有氧氧化肌糖原、脂肪酸、酮体
- 剧烈运动时:无氧氧化
- 乳酸循环:是整合糖异生与肌糖酵解途径的重要机制
- 不同骨骼肌产能方式不同
- 脂肪组织
- 摄取的能量存储于脂肪组织
- 膳食脂肪:以乳糜微粒的形式运输至脂肪,在脂蛋白脂肪酶的作用下被水解摄取,用于合成脂肪细胞内脂肪存储
- 膳食糖
- 运输至肝转化为脂肪,以VLDL的形式运输至脂肪组织,在LPL作用下被水解摄取,合成脂肪存储于脂肪细胞
- 脂肪细胞也能将糖转化为脂肪储存
- 饥饿时主要靠分解存储于脂肪组织的脂肪供能
- 脂解激素将脂肪组织的能量以脂肪酸和甘油的形式释放入血
- 肝可以将脂肪酸分解为酮体,经血液运输至肝外利用
- 摄取的能量存储于脂肪组织
- 肾
- 供能
- 肾髓质无线粒体,主要靠糖酵解供能
- 肾皮质主要靠脂肪酸及酮体氧化供能
- 糖异生:
- 一般情况下肾糖异生产生的葡萄糖较少,只有肝的 10%,但长期饥饿后,糖异生的量大量增加,可跟肝几乎相等
- 肾糖异生有调节酸碱平衡的作用
- 酮体生成:极少
- 供能